- Review
- Open access
- Published:
- Ankush Yadav1,
- Suhani Sharma1,
- Nitesh1,
- Rinku Meena1,
- Rupesh Bhardwaj1,
- Prashant Swapnil1 &
- …
- Mukesh Meena2
Biotechnology for Biofuels and Bioproducts volume 18, Article number: 92 (2025) Cite this article
Abstract
Population growth throughout the world has led to increased pollution and overconsumption of fossil resources. Microalgae are increasingly recognized as sustainable biofactories for producing lipids and astaxanthin, two commercially significant metabolites with wide-ranging applications in biofuel, pharmaceutical, cosmetic, and nutraceutical industries. Enhancing the yields of these compounds remains a major challenge due to growth–productivity trade-offs and limited understanding of regulatory mechanisms. This review aims to bridge that gap by providing a comprehensive and comparative analysis of traditional and modern strategies employed to enhance lipid and astaxanthin production in microalgae. We critically evaluate stress-based methods (e.g., salinity, light, nutrient limitation), phytohormone treatments, cultivation system optimization, and genome editing technologies, including CRISPR/Cas9. Special emphasis is given to gene-level responses and pathway-level regulation involved in these enhancements. This review article highlights the novel synchronization between astaxanthin and fatty acid biosynthesis under various stress conditions which emphasizes the role of diacylglycerol acyltransferase (DGAT) enzymes to enhance astaxanthin accumulation. Editing technologies with base suggest a novel strategy to reduce off-target effects and enhance metabolic efficiency related to lipid and astaxanthin biosynthesis.
Graphical Abstract

Introduction
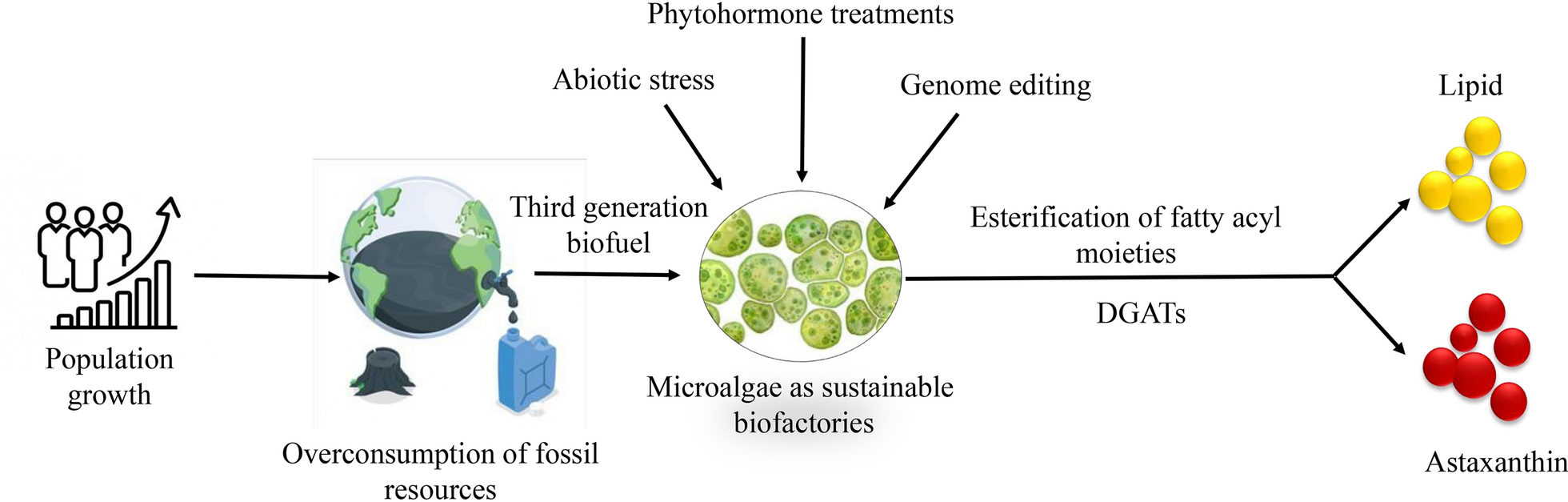
According to United Nations current world’s population is expected to increase by nearly 2 billion persons in the next 30 years, from the current 8–9.7 billion in 2050, which is the main reason for the overconsumption of fossil fuels which ultimately leads to different kinds of pollution. Biofuels have witnessed significant advancements in recent years, with different generations of raw organic materials being utilized for production (Fig. 1). First-generation biofuels are made from feedstocks that are also suitable for human consumption, such as maize, sugarcane, palm oil, sugar beetroot, and wheat [1]. However, non-edible food crop components, including stems, leaves, and husks, are used to create second-generation biofuels, which offer an alternative to waste utilization [2]. Despite their potential, these generations of biofuels face limitations as they compete with food crop production and require cultivable land, which is a finite resource. Third-generation biofuels by microalgae, which convert carbon dioxide (CO2) and light into biomass and high-energy lipids, are used to develop solutions to these problems [3]. Biofuel production from microalgae is an important area of research due to concerns over the negative impact of the overuse of fossil fuels on the environment, because of their non-renewable nature, harmful after-use products and high global demand [4]. Compared to previous generations, third-generation biofuels offer several advantages, including reduced competition for food crops and available farmland, lower water requirements, and higher CO2 mitigation rates [5]. Additionally, compared to second-generation biofuels made from oleaginous crops, microalgae biofuels exhibit greater carbon absorption and lipid content, up to 15–20 times higher [6, 7].

Different generations of biofuels with their respective methods
Biotechnology has led to the expansion of fourth-generation biofuels, utilizing gene editing technology in microalgae, to produce bioenergy efficiently. Notably, microalgae exhibit rapid growth rates under favorable conditions, enabling higher biomass production rates than land crops. Microalgae provide over 40% of the world’s carbon fixation; therefore, using them to produce biofuels offers a big chance to cut greenhouse gas emissions [8]. Additionally, certain microalgae strains have been shown to contain as much as 70% oil by dry weight, indicating their potential as a significant source of biofuels. By using microalgae’s special qualities and capacities, we can get past the drawbacks of earlier biofuel generations and promote a more economically and environmentally sound energy source. The cultivation of microalgae offers a sustainable pathway towards reducing our dependence on fossil fuels and mitigating the detrimental effects of greenhouse gas emissions. Biofuels have become a viable substitute for fossil fuels in many regions of the world, including the United States, Europe, Asia, and Brazil [9]. Till now, these parts of the world mainly use sugarcane, palm oil, and vegetable oil waste to produce biofuels. However, the unique properties of microalgae, which capture carbon and incorporate it into metabolism, attract researchers to the production of biofuels. Biofuels can be used to control pollution and provide various job opportunities and income growth for farmers [10]. However, a poor biorefinery system is one of the major drawbacks of microalgal biofuel production. The high cost of the biorefinery of microalgal biofuels makes it an unprofitable approach. Combining biofuels with different byproducts might be a good way to reduce costs [11]. Microalgae such as Haematococcus pluvialis produce various kinds of secondary metabolites (astaxanthin, polyphenols, vitamins, fatty acids, and phycobiliproteins) and offer significant applications as antimicrobial agents, antioxidants, nutritional supplements, and colorants under various stress conditions [12, 13]. As a carotenoid, astaxanthin is derived from its precursor molecule, β-carotene [14, 15]. It undergoes a series of enzymatic reactions, including oxidation and cyclization, to form astaxanthin. This conversion occurs within the cells of specific organisms, primarily microalgae and some bacteria [16]. Concurrent fatty acids and astaxanthin production in microalgae presents a highly efficient and economically viable approach for sustainable bioproduct manufacturing. Both metabolites share common precursors such as acetyl-CoA and are regulated by similar stress-responsive pathways, which allows for simultaneous enhancement under optimized conditions [17]. Physiological stressors like nutrient limitation, high light intensity, and salinity can trigger lipid accumulation while stimulating astaxanthin biosynthesis, making it possible to harness both high-value compounds in a single cultivation cycle.
The antioxidant activity of astaxanthin is well-known and much greater than that of various other carotenoids, including lycopene and β-carotene [18]. Because of its special molecular makeup, it can efficiently moderate reactive oxygen species as well as eliminate radicals, shielding the cells from oxidative damage. This antioxidant capacity is particularly relevant in the context of medicine, where oxidative stress is implicated in various diseases and ageing processes [19]. Astaxanthin has drawn interest from the medical community due to its possible health advantages. It has been studied extensively for its antioxidant properties and its function in lowering oxidative stress, inflammation, DNA damage [20], mitigating age-related macular degeneration [21], reduce the expression of inflammatory enzymes [22], protective effects against cardiovascular diseases by reducing oxidative stress [23], improving lipid profiles [24], and enhancing vascular function (Fig. 2). While astaxanthin is naturally present in certain organisms, it can also be produced synthetically or through microalgal culture.

Biomedical applications of astaxanthin
The cost of synthesis of astaxanthin by microalgal culture varies based on the particular strain of microalgae, conditions for cultivation, and techniques for extraction. However, astaxanthin production through microalgae cultivation is generally considered more sustainable and environmentally friendly than synthetic methods. These byproducts have high market value and can be co-cultivated with biofuels and act as an additional source of income. Microalgae may be used to create a variety of biofuels, including bioethanol, biodiesel, bio-oil, biohydrogen, biogas, and charcoal [25]. However, most of the green microalgae produce lipids in high amounts as compared to carbohydrates in their cells, making the multiple biofuel production unrealistic [26]. To overcome this limitation, advanced microalgae strains can be used for biofuel production. When vital elements like sulfur or nitrogen are depleted in microalgae, cell division is inhibited and fat buildup is set off [27]. The continuous fixation of carbon, coupled with restricted growth, diverts metabolic pathways towards lipid synthesis. So, it is concluded that microalgae with a fast growth rate possess very low lipid content [28]. In order to overcome this distinct type of physiological stress, a molecular strategy and dietary stress have been employed to enhance the lipid output of microalgae [29].
Till now, these approaches have been successfully used for increasing the productivity of lipids, β-carotene, and carbohydrates. Genetic engineering gained more attention from these approaches with the advancement of tools and resources, and it is easy to manipulate the genome more accurately. The molecular approaches, like the development of CRISPR/Cas9 (clustered regularly interspaced short palindromic repeats) tool in recent years, give efficient ways of genetic manipulation in microalgae, generally involving several steps, regardless of the specific technique employed [30]. Host selection is crucial, as different microalgae species possess varying metabolic properties and growth characteristics. The intended metabolic route and the particular application determine which host organism is suitable. Finding the target gene is necessary after choosing the host organism. This necessitates a deep comprehension of the genes regulating the pathway of metabolism being altered. By selecting the appropriate gene target, researchers can effectively modulate the metabolic processes in microalgae. Plasmid construction plays a vital role in introducing foreign DNA into microalgae. The plasmid serves as a vehicle for delivering the desired genetic material into the host organism. It typically contains the gene of interest, along with regulatory elements that control its expression. Constructing plasmids that are compatible with the microalgal system is essential for successful transformation. The selection of transformation tools is another critical aspect of genetic manipulation in microalgae. Different methods, such as electroporation, particle bombardment, or Agrobacterium-mediated transformation, can be employed depending on the specific requirements of the experiment. The choice of transformation tool affects the efficiency and ease of introducing the desired genetic material into the microalgal cells.
Recent reports discussed that phytohormones significantly increase lipid production in microalgae [31] by regulating biosynthesis genes and modulating reactive oxygen species (ROS) levels under stress conditions [32]. Key enzymes involved in fatty acid biosynthesis are overexpressed to enhance lipid content through salicylic acid [33]. Finally, DNA editing tools, such as CRISPR/Cas9, enable precise modifications of the microalgal genome. With high efficiency and accuracy, these tools allow researchers to introduce targeted genetic changes, such as gene knockout, gene insertion, or point mutations. By harnessing the power of DNA editing tools, scientists can achieve specific metabolic modifications in microalgae. The fundamental steps of any genetic manipulation technique are host selection, target gene, plasmid production, transformation tools selection system, and DNA editing tools.
Lipids in microalgae
Microalgae are unicellular photosynthetic microorganisms that produce a large amount of lipids and have been considered as a good source for biofuel, nutraceuticals, and pharmaceuticals [34]. Various types of lipids such as triacylglycerols (TAGs), glycolipids, phospholipids, and sterols, were produced by microalgae by utilizing a suitable comprehensive database and analytical methods. TAGs serve as the primary storage lipids in microalgae, and they account for up to 50–80% of total lipid content [35]. Sulfoquinovosyldiacylglycerol (SQDG) and monogalactosyldiacylglycerol (MGDG) are two glycolipids that are crucial for photosynthesis and are significant parts of the thylakoid membrane in the chloroplast [36]. Cell membrane lipids like phosphatidylcholine (PC) and phosphatidylethanolamine (PE) work as key building blocks in microalgae [37]. The fluidity of the cell membrane is mainly regulated by sterols, including cholesterol and ergosterol, which are significant parts of the cell membrane [38]. Lipid biosynthesis is a multifaceted process that takes place in several routes.
De novo synthesis of lipids in microalgae
Microalgal lipids offer an opportunity to reduce the impact of global energy demand for fuels. The systems biology approach elucidates the key components for the commercialization of microalgal-derived biofuels from oleaginous microalgae by initiating strain-improvement strategies [39]. Photosynthesis plays a pivotal role in lipid biosynthesis by converting CO2 into glycerate-3-phosphate (G3P). G3P serves as a crucial precursor molecule for the synthesis of various storage materials, including polysaccharides and lipids. A crucial first step in lipid biosynthesis is converting G3P to pyruvate, which is then converted to acetyl-CoA by the pyruvate dehydrogenase complex (PDC) [40]. This conversion occurs within the plastid, the organelle where lipid biosynthesis predominantly occurs. Acetyl-CoA, the product of G3P metabolism via the PDC, is a vital building block for lipid synthesis. It serves as a central precursor molecule that contributes to producing fatty acids, glycerol, and other lipid components. Furthermore, another metabolic process can produce acetyl-CoA, allowing polysaccharides to be converted into lipids [41]. This alternative pathway supports lipid biosynthesis by providing an extra mechanism for generating acetyl-CoA, which is frequently used by oleaginous heterotrophs during sugar absorption [42].
The ability of photosynthesis to convert CO2 into G3P represents a crucial link between carbon fixation and lipid biosynthesis. Wang et al. [43] explored the improvement of lipid production via G3P synthesis photosynthetically. By harnessing energy from sunlight and utilizing CO2, photosynthesis provides the necessary carbon and energy sources to drive the synthesis of lipids. This interplay between photosynthesis and lipid biosynthesis highlights the fundamental role of plants, algae, and other photosynthetic microorganisms in producing and storing lipids, which are essential for energy storage, structural integrity, and cellular signaling. De novo synthesis of TAGs takes place in chloroplast green microalgae (Fig. 3). The first important step in this process is the conversion of acetyl-CoA into malonyl-CoA, which is catalyzed by the enzyme acetyl-CoA carboxylase (ACCase) [44], which acts as the enzyme responsible for slowing down the production of fatty acids. This step ensures the availability of malonyl-CoA, which is essential for subsequent fatty acid synthesis [45]. After malonyl-CoA is formed, an enzyme called malonyl-CoA transacylase transforms it into malonyl-ACP (acyl carrier protein) serving as an essential intermediate in the elongation of acyl chains during fatty acid synthesis [46].

De novo biosynthesis of TAG in green microalgae. Enzymes are represented in red. ACCase: acetyl-CoA carboxylase; ACP: acyl carrier protein; DAG: diacylglycerol; DGAT: diacylglycerol acyltransferase; DGD: digalactosyldiacylglycerol synthase; FAS: fatty acid synthase; FAT: fatty acyl-ACP; G3P: glycerol 3 phosphate; GPAT: glycerol-3-phosphate acyltransferase; LPA: lysophosphatidic acid; LPAT: lysophosphatidic acid acyltransferase; MCMT: malonyl-CoA:acyl carrier protein malonyltransferase; MGD: monogalactosyldiacylglycerol synthase; MGDG: monogalactosyldiacylglycerol; PA: phosphatidic acid; PAP: phosphatidic acid phosphatase; PDH: pyruvate dehydrogenase
Fatty acid extension and desaturation pathway
Malonyl-ACP serves as an important intermediate in the elongation of acyl chains during fatty acid synthesis. The elongation process is subsequently catalyzed by the enzyme fatty acid synthase (FAS), which uses acetyl-CoA and malonyl-ACP as substrates [47], and fatty acid chains gradually extend. Different fatty-ACP thioesterases (FAT) promote the cessation of fatty acid chain elongation. These thioesterases are essential for catalyzing the escape of unbound free fatty acids (FFAs) from acyl-ACP by hydrolyzing it [48]. This step is significant as it contributes to the generation of FFAs that can subsequently be utilized for the synthesis of TAGs.
Polyketide synthase (PKS) pathway
Apart from the FAS pathway, some microalgae also possess a polyketide synthase (PKS) pathway, which functions as an alternative mechanism for lipid biosynthesis. Unlike the FAS system, which relies on a modular enzymatic assembly line, the PKS pathway involves iterative catalysis by multifunctional enzymes [49]. PKS is an oxygen-independent pathway to synthesize unsaturated fatty acids and is limited to only some families, unlike n-3 and n-6, which are widely represented among microalgae taxa. It has similarities with the FAS pathway as both need four basic reactions and the same enzymes [50]. This pathway enables the biosynthesis of complex lipids and polyketides with unique structural variations. The PKS pathway can be particularly significant in certain microalgae, which contributes to the diversity of lipid products and potentially enhances the production of high-value lipids for biofuels and nutraceuticals. The FAS pathway requires much energy, while the PKS pathway is more efficient at producing polyunsaturated fatty acids (PUFAs) with fewer steps. Although typically anaerobic, PKS can also work in aerobic conditions [49].
TAG biosynthesis and assembly
TAGs are synthesized through a series of enzymatic reactions using G3P as the basic carbon substrate. The first step involves glycerol-3-phosphate acyltransferase (GPAT) acylating G3P at the sn-1 position to produce lysophosphatidate (LPA). Lysophosphatidate acyltransferase then adds a fatty acid to the sn-2 position of LPA to produce phosphatidate (PA). Phosphatidate acid phosphatase (PAP) removes the phosphate group from PA’s sn-3 position and produces diacylglycerol (DAG). TAG is created when diacylglycerol acyltransferase (DGAT) acylates DAG at its sn-3 location to complete the process [51]. Fan et al. [52] stated that in N-starved Chlamydomonas, TAG production depends on de novo fatty acid production in the chloroplast. They found that the type-2 fatty acid synthase-specific inhibitor cerulenin significantly inhibited the buildup of TAG brought on by N deprivation. This suggests that during food scarcity situations, fatty acid synthesis plays a crucial role in the formation of TAG.
Lipids as biofuel
TAGs are essential for microalgae cells’ ability to store energy. Through a convoluted process, glycerate-3P is transformed into molecules like pyruvate, sugar, xylose, acetate, amino acids, and triglycerides that are essential for the metabolism, development, storage of energy, and maintenance of cells. The processing of TAGs into fatty acid methyl esters, a vital and adaptable form of biodiesel and the basis for its production, is simple and may be accomplished by the trans-esterification process [53]. The effective trans-esterification process requires catalysts (alkaline and acidic) for a low-cost, less corrosive, and environmentally friendly approach [54]. Worldwide biodiesel production currently revolves around using alkaline and acidic catalysts employed in heterogeneous and homogeneous phases. The composition of the lipids used significantly impacts the production process and the quality of the resulting biodiesel. In contrast to polar lipids like phospholipids and glycolipids, which are undesirable due to their detrimental effects on emulsification and catalyst depletion, TAGs are the ideal raw materials for manufacturing biodiesel. Additionally, non-TAG lipids can contribute to higher sulfur and phosphorus content, which can adversely affect fuel quality [55].
The degree of unsaturation of fatty acids (FAs) is critical in determining how well biodiesel performs overall. Since the inclusion of PUFAs might decrease the stability of the biodiesel, the main constituents of biodiesel are saturated fatty acids (SFAs) and monounsaturated fatty acids (MUFAs) [56]. Polyunsaturated fatty acids (PUFAs), such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), enhance microalgal lipid yield and improve the cold flow properties of biodiesel, making it more suitable for low-temperature applications. However, their high unsaturation level increases susceptibility to oxidation, shortening the biodiesel’s shelf life. Additionally, the lower cetane number of PUFAs results in incomplete combustion and higher emissions. They also have lower energy content and involve high processing costs [57, 58].
Techniques for improving lipid content in microalgae
Different types of physiological stresses and molecular approaches can significantly impact lipid enhancement strategies in microalgae (Fig. 4). These stresses can affect the growth, metabolism, and overall lipid production of microalgae, ultimately influencing the efficiency of lipid enhancement processes (Table 1). Here are the general effects of physiological stresses and molecular approaches on lipid enhancement strategies in microalgae.

Different types of physiological stresses and molecular approaches that can significantly impact lipid enhancement strategies in microalgae
Light
Light intensity affects photosynthesis, which consequently affects the formation of ROS during photosynthetic electron transport. Light is essential for the formation of lipids in microalgae, which significantly affects biomass output. One important component influencing the lipid synthesis process is light wavelength, which affects the microalgae’s ability to produce lipids in different ways. It has been demonstrated that blue light with a wavelength of around 450–470 nm increases the lipid accumulation in microalgae [68]. Studies show that microalgae exposed to blue light, such as Chlorella vulgaris [69] or Nannochloropsis sp. [70], significantly increased lipid content. Moreover, red light in the 640–680 nm range has also been found to stimulate lipid production in microalgae. Studies have shown that red light increases the activity level of certain important lipid biosynthesis-related enzymes, including fatty acid synthase and acetyl-CoA carboxylase. These enzymes play crucial roles in converting CO2 and light energy into lipids. Interestingly, the combination of red and blue light has shown remarkable results in boosting lipid production in microalgae. When exposed to a combination of these two wavelengths, microalgae species like H. pluvialis [71] and Scenedesmus obliquus [72] have exhibited a substantial increase in lipid content. For instance, studies have reported a lipid increase of up to 50% in H. pluvialis when subjected to red and blue light combined [68].
Red and blue light work in concert to maximize the photosynthesis rate of microalgae by increasing light absorption and promoting the activation of important lipid biosynthesis-related enzymes [69]. The particular mix and length of red and blue light exposure may change based on the kind of microalgae and the intended results for lipid formation. The time duration of the light/dark period plays a crucial role in the lipid biosynthesis of microalgae, influencing the composition of saturated and unsaturated fatty acids [68]. During the dark period, microalgae rely on stored carbohydrates as a source of energy through respiration, leading to a decrease in lipid accumulation, particularly in saturated fatty acids. Cha et al. [73] stated that Chlorella vulgaris cultures subjected to a dark period showed a reduction in saturated fatty acids, including palmitic acid (C16:0). Conversely, exposure to light stimulates photosynthesis in microalgae, driving lipid synthesis and subsequent accumulation. Numerous studies have demonstrated that the generation of lipids is also significantly impacted by light intensity. Cheirsilp et al. [74] studied the effect of light intensity on the amount of lipids accumulated in Nannochloropsis sp. and discovered that increased light intensity led to higher lipid content. They observed that when the illumination intensity was raised from 50 to 200 μmol photons/m2/s1, the lipid content increased by 50%. Different microalgal species exhibit varying responses to light intensity, with some species showing optimal lipid accumulation at specific light intensities. In contrast to lower light intensities research by Nzayisenga et al. [75], Scenedesmus obliquus showed a 72% rise in lipid content when subjected to 120 μmol photons/m2/s1.
This study highlighted the species’ maximum lipid content. In terms of culture methods, the kind of system used, such as open pond systems or photobioreactors, can also impact how light affects lipid production [76]. Lipid synthesis can be impacted by the amount, quality and spectrum of sunlight as well as the possibility of light attenuation from things like cloud cover. However, because of the closed systems, photobioreactors provide more control over the level of light, time span, and spectrum makeup. This control enables optimization of light parameters for enhanced lipid accumulation in specific microalgal species.
Carbon dioxide (CO2)
Elevated atmospheric CO2 concentrations result from human-induced processes such as the combustion of fossil fuels and deforestation, which contribute to the ongoing increase in CO2 emissions. Microalgae use CO2 as a vital source of carbon. They may use this inorganic carbon to create biomass and interesting chemical compounds when exposed to sunlight. There is a specific maximum and minimum level of CO2 for the effective growth of microalgae. Makareviciene et al. [77] found that microalgae exhibit a lower growth rate and reduced lipid production when exposed to suboptimal CO2 concentrations. The study identified a minimum CO2 level of 0.04% (v/v) for sustained microalgal growth. Below this threshold, photosynthesis is limited, leading to decreased biomass accumulation and lipid productivity. While microalgae require CO2 for growth, excessively high concentrations can have detrimental effects.
Cheng et al. [78] demonstrated that elevated CO2 levels (>10% v/v) can inhibit microalgal growth due to reduced light penetration and increased pH. These extreme CO2 conditions negatively impact photosynthesis, leading to cellular damage and reduced lipid biosynthesis. Under high CO2 concentrations, microalgae can actively uptake CO2 via carbonic anhydrase enzymes [79]. Following this absorption, a sequence of chemical processes converts CO2 into bicarbonate (HCO3‾). While HCO3‾ provides an alternative carbon source, its accumulation disrupts various metabolic reactions critical for lipid biosynthesis [80]. Microalgae employ carbon concentrating mechanisms (CCMs) to enhance CO2 uptake and concentration within cells. However, excessive CO2 availability can also disrupt these CCMs, impairing the efficient utilization of CO2 for lipid synthesis. The effectiveness of CO2 absorption and assimilation can be decreased by downregulating CCM-related genes, as reported by Barati et al. [81] at high levels of carbon dioxide (>5% v/v). High levels of CO2 cause microalgae to divert carbon flow towards the synthesis of carbohydrates rather than lipids [46]. The increased activity of enzymes that participate in carbohydrate metabolism, such as ADP-glucose pyrophosphorylase and starch synthase, which increase the manufacture of starch as a carbon storage molecule, is thought to be the cause of this change in metabolic pathways.
Furthermore, the enzymes controlling the lipid production may be inhibited by the excess HCO3− those forms at high CO2 concentrations. Acetyl-CoA carboxylase and fatty acid synthase are two important enzymes in the lipid biosynthesis pathway whose function is adversely affected by high HCO3− levels [82]. This disruption hampers the conversion of acetyl-CoA into fatty acids, resulting in decreased lipid production. Moreover, it has been discovered that elevated CO2 concentrations modify the regulation of genes related to lipid metabolism. To prove this, these studies conducted by Zhang et al. [83] demonstrated that elevated CO2 levels upregulate genes related to lipid degradation and down-regulate genes involved in lipid production in microalgae. This deregulation of gene expression further contributes to the reduction in lipid accumulation. The disturbance of metabolic pathways under high CO2 concentrations is not limited to lipid biosynthesis, it can also disrupt the balance between carbon and nitrogen metabolism in microalgae [84]. The optimum conditions for the maximum biomass and lipid surface productivity were 28.0 ± 1.5 g/m2/day at 730 μE/m2/s with 8% and 4.2 ± 0.3 g/m2/day at 500 µE/m2/s with 7% of CO2 concentration [85].
Temperature
Lipid accumulation in microalgae is greatly influenced by temperature and also affects the algal development, metabolism, and fatty acid composition [86]. Understanding the effects of temperature is crucial for optimizing lipid production in microalgae-based biofuel production systems. Firstly, microalgae exhibit an optimal temperature range for lipid accumulation. Different species have specific temperature requirements for optimal growth and lipid production. Ferrer-Ledo et al. [87] stated that the microalga Nannochloropsis oceanica showed the highest lipid content at 25 °C, while lower and higher temperatures reduced lipid accumulation. Chlorella vulgaris, a widely studied microalga, showed enhanced lipid content up to 35 °C, beyond which lipid accumulation declines [88]. However, the halotolerant microalga Scenedesmus strain shows maximal lipid synthesis at temperatures of 30 °C [89]. These illustrations showed how temperature preferences for lipid formation in microalgae are species-specific. Understanding and controlling the temperature conditions are vital for maximizing lipid productivity in different microalgal species. This indicates that maintaining the appropriate temperature is essential for maximizing lipid productivity and affects the activity and stability of enzymes that regulate lipid production in microalgae.
Certain enzymes have been reported which are responsible for fatty acid biosynthesis and sensitive to temperature variations. These variations can influence the lipid accumulation process. Leyva et al. [90] examined that temperature affected acetyl-CoA carboxylase (ACC), a crucial enzyme for fatty acid production in Chlorella vulgaris, and resulted in ACC activity increased with temperature up to an optimal range and then declined. Moreover, changes in temperature can impact the fatty acid content of microalgae, particularly the proportion of polyunsaturated fatty acids. PUFAs, including omega-3 and omega-6 fatty acids, are valuable in nutrition and industry. The variation in temperature resulted in an increase of fatty acid composition (total percentage of PUFAs) of Nannochloropsis spp., particularly EPA [91]. This study suggested that alteration in temperature can be used as a strategy to enhance PUFA content in microalgae for various applications in nutraceuticals and functional foods.
Nutrient stress
Various studies have indicated that any kind of modification in nutrient contents, such as phosphorus (P), salinity, and nitrogen (N), can play a beneficial role in lipid synthesis [92]. These nutrients are crucial in vital cellular mechanisms such as photosynthesis, protein synthesis, and cell division in microalgae. In nutrient deficiency or limitation conditions, microalgae utilize their energy on cellular defence against oxidative stress, which ultimately results in the production of neutral lipids and other valuable byproducts like carotenoids, phycocyanin, eicosapentaenoic acid, and docosahexaenoic acid [93]. Encouraging microalgae to prioritize lipid accumulation by adjusting the conditions of nutritional stress is feasible, which creates opportunities for increased lipid synthesis in microalgal growing systems.
Nitrogen
For microalgae, nitrogen is a necessary ingredient involved in many cellular functions such as cell division, protein synthesis, and the chlorophyll synthesis. Assimilation of nitrate is crucial for plant and microorganism growth [94]. However, when microalgae are subjected to nitrogen deficiency, their metabolic pathways undergo significant changes, which lead to an enhancement of lipid synthesis. Nitrogen deficiency in microalgae triggers a series of physiological responses aimed at adapting to the limited nutrient availability. One of the primary effects is the inhibition of cell division, as nitrogen is crucial for DNA replication and cell proliferation [95]. Microalgae prioritize survival over growth, leading to a halt in cell division and an accumulation of biomass. As the cell division process slows down, microalgae redirect their metabolic resources towards alternative pathways, including lipid synthesis [96]. Under nitrogen deficiency, microalgae often store more energy and sequester carbon by accumulating bigger amounts of neutral lipids like TAGs. Intricate regulatory mechanisms govern the shift towards lipid synthesis in microalgae under nitrogen deficiency. To increase the synthesis and formation of lipids, many important enzymes involved in lipid biosynthesis, including FAS, ACC, and DGAT, are upregulated [97]. In addition, a nitrogen shortage causes microalgae to have higher total lipid content as well as a different fatty acid distribution.
In certain species, nitrogen deprivation can lead to a significant increase in desirable fatty acids, including PUFAs such as EPA and DHA. This change in fatty acid composition adds value to microalgal lipids for nutraceuticals and biodiesel production applications. In nitrogen-limited circumstances, Chlorella vulgaris and Nannochloropsis sp. showed approximately 2 to 5 and 7- to tenfold increments in lipid content, respectively [98, 99]. This substantial enhancement in lipid accumulation showcases the species’ adaptive response to nitrogen stress. Furthermore, Scenedesmus obliquus showed a 40–60% increment in lipid content under nitrogen-deprived conditions [100]. These examples highlight the species-specific character of these reactions and highlight the possibility of nitrogen deprivation as a tactic for promoting lipid formation in microalgae.
Heavy metal stress
Metals such as Cu (copper), Fe (iron), Mn (manganese), Ni (nickel), and Zn (zinc) are vital in microalgal cells as precursors of vitamins, catalytic cofactors, and key components of metalloenzymes and cell membrane structural proteins. Copper participates in electron transport and enzyme activities, while iron is essential for chlorophyll synthesis, electron transfer, and biomolecule production. Manganese acts as a cofactor in redox reactions and protects against oxidative stress. Nickel is crucial for nitrogen metabolism and enzymatic processes, and zinc is required for enzyme activity, protein synthesis, and membrane integrity. These metals play critical roles in microalgal cellular functions, spanning photosynthesis, respiration, and metabolic pathways. But some metals, such as Cd (cadmium), Cr (chromium), Pb (lead), Hg (mercury), and As (arsenic), can be harmful to algal cells, affecting their ability to photosynthesize, preventing cell division, and reducing enzyme activity [101].
Heavy metals induce the production of ROS, causing oxidative stress and damaging cellular components. Chlorophyll synthesis is inhibited by heavy metals, impairing photosynthesis and reducing energy conversion. Enzyme activities and metabolic pathways are disrupted, hindering vital cellular processes. Heavy metals can also disturb cell proliferation and growth by interfering with the cell cycle. Additionally, they damage the cell membrane, compromising its integrity and permeability. Microalgae have garnered attention for their potential in bioremediation, specifically in removing and sequestering heavy metals from polluted environments. They possess polyphosphates, phytochelatins, and metallothioneins to capture and detoxify heavy metals, reducing their adverse impacts on ecosystems. Polyphosphates, high-energy phosphate compounds accumulated within microalgal cells, bind and sequester heavy metals, reducing their bioavailability and toxicity [102]. Small peptides called phytochelatins are produced in the wake of stress caused by heavy metals and combine with heavy metals to create stable complexes that help in their detoxification [103]. Metallothioneins, small metal-binding proteins produced by microalgae, protect against heavy metal toxicity by binding and sequestering them [104].
In different studies, it is found that heavy metal stress can also enhance the lipid productivity in microalgae. An accumulation of different metals, such as Fe2+, Fe3+, Ca2+, Mg2+, and Co2+, has varying effects on the lipid productivity of microalgae. According to Ren et al. [105], the lipid profile of Scenedesmus sp. increased from 9 to 13% when the Fe3+ concentration was raised from 0 to 0.1 g/L. For instance, microalgae subjected to 20 mg/L Mg2+ showed nearly a fourfold rise in biomass and a 1.1-fold rise in the production of lipids compared to the control [106]. Furthermore, it has been discovered that appropriate Ca2+ and Cu2+ concentrations encourage microalgae proliferation and lipids’ synthesis. In H. pluvialis, after 15 days of culture, the total lipid levels attained their maximal levels (3.32%) under 6 μM Cu treatments, which increased by 34.99% with the non-treated sample [107]. When microalgae Nannochloropsis oculata was treated with 4.6 mg/L Cu2+ 77.1% lipid content was increased as compared to the control [108].
According to Sun et al. [109], arsenic caused the lipid profile of Nannochloropsis sp. to rise at 10 mg/L of arsenic concentration, and the highest level of 24.2% was recorded. Similarly, algal cells that were subjected to 80 and 40 μM of Cd2+ showed 160 and 140% increments in lipid content [110]. One possible explanation for the increased lipid synthesis might be the generation of antioxidants, which can mitigate the oxidative damage brought on by exposure to Cd2+. Yang et al. [111] investigated lipid accumulation in Chlorella minutissima UTEX2341 impacted by Cd2+ and Cu2+. They discovered that Cd2+ and Cu2+ significantly influenced the buildup of triglycerides under heterotrophic cultivation conditions. At 0.4 mM of Cd2+ and 0.4 mM of Cu2+, the maximum yield of lipids was recorded, attaining 249.4, and 154.2 mg/L/d, respectively. León-Vaz et al. [112] reported a 560% rise in plastid acyl-ACP desaturase under Cd2+ stress, which enhances lipid accumulation.
Additionally, when Chlorella sorokiniana was exposed to Cd2+, it showed increased expression of antioxidant enzymes, photorespiration, and carbohydrate catabolism. They discovered that Cr (III) improved the resistance to oxidation of biodiesel by increasing the level of methyl oleate (C18:1) and lowering the contents of methyl linolenate (C18:3) and methyl linolelaidate (C19:1). Microalgae demonstrated a remarkable 99% retention effectiveness for 100 mg/L Cr (III), most likely as a result of their capacity to accumulate lipids and metabolites in the presence of heavy metals, hence mitigating the effects of oxidative damage. These results demonstrate the capacity of heavy metals like As, Cd, and Cr to encourage the accumulation of lipids in microalgae and the consequences this has for improving the resistance to oxidation and producing biodiesel. Battah et al. [113] investigated the effects of different concentrations of MnCl2 at 2, 10, and 12 μM on C. vulgaris lipid composition, they found substantial rises of 14, 16, and 15%, respectively. Additionally, they discovered that, in comparison to the control, including cobalt nitrate at various doses increased the lipid content by an extra 25%. In a different investigation, it was shown that when C. vulgaris was cultivated with different Fe3+ concentrations, there was an astounding 56.6% rise in total lipid levels.
Salinity stress
Salts are vital elements that affect numerous metabolic processes and are crucial to the growth and development of microalgae. In addition to their role in maintaining osmotic balance, salts are involved in enzyme activation, ion transport, and protein synthesis [114]. Understanding the impact of salt stress on lipid accumulation in microalgae is of major significance for optimizing lipid synthesis and improving the feasibility of microalgae as a sustainable source of biofuel [115]. Salt stress can trigger various physiological and biochemical responses in microalgae as well as cyanobacteria [116], leading to enhanced lipid accumulation. Gour et al. [117] performed experiments on three different microalgal species, Scenedesmus quadricauda (Sq19), S. dimorphus (Sd12), and Chlorella sp. (Chl16), with a salinity level of 40 mM NaCl in BG-11 medium, the maximum specific rate of development and daily doubling were recorded. Furthermore, on BG-11 medium, the corresponding algal isolates were able to attain maximal lipid content and lipid output at 160 mM NaCl. For Sq19, Sd12, and Chl16, the maximal lipid content was measured at 18.28, 30.70, and 32.19% while the lipid productivity reached 8.59, 13.81, and 10.27 mg/L/d, respectively.
Additionally, the development of Sd12 was stimulated by a stepwise salinity stress of 160 mM NaCl, which led to greater lipid content (39.42%) than the control group (20.4% lipid content). Under NaCl stress development and buildup of lipids were found in S. quadricauda FACHB-1297 [118]. The findings demonstrated that in comparison to the BG-11 medium, biomass concentration and lipid content were considerably greater (2.15–2.68 times) at NaCl concentrations ranging from 0.88 to 5.85 g/L. Under 2.63 g/L NaCl concentration the greatest lipid content, biomass level, total nitrogen elimination rate, total phosphorus reduction effectiveness, sugar content, and protein content were found 39.33, 0.72, 39, 100, 19.26, and 5.38%, respectively.
Bartley et al. [119] investigated the effect of salt stress on N. salina and found lipid increment at 34 PSU (particle salinity unit). According to Salama et al. [120] at 25 mM NaCl Chlamydomonas Mexicana and S. obliquus, achieved maximum lipid content of 37 and 34%, respectively. Pandit et al. [121] examined C. vulgaris and Acutodesmus obliquus, reporting maximum lipid content of 49 and 43%, respectively, at 0.4 M NaCl. A. dimorphus showed significant lipid accumulation (33.40 ± 2.29%) at 200 mM NaCl, increasing to 43% with extended saline stress.
Phytohormones induction for lipid production enhancement
Environmental stressors help in the accumulation of secondary metabolites but suppress cell growth. Phytohormones are proving to play an important role in resolving this problem. They can regulate and control the expansion of microalgal cell growth [122]. The approach to using phytohormones is becoming popular to manipulate the microalgal metabolism and produce desired secondary metabolites. Phytohormones are reported to regulate cell growth, secondary metabolism and stress tolerance in microalgae. Phytomelatonin, has been found to improve stress resistance and enhance the synthesis of metabolites by maintaining redox balance and regulating the expression of genes involved in metabolite synthesis. Exogenous melatonin application increased lipid content as well as biomass concentration, under nitrogen deficiency in Chlamydomonas reinhardtii [123]. Melatonin has improved the lipid content and astaxanthin production. In a study on the combined effect of melatonin (10 µM) with red LED stimulated in microalgal growth resulting in 200% biomass productivity and 94% lipid content increase in C. vulgaris cultured in dairy waste [124]. Similarly, the application of 32 µM linoleic acid under light stress enhanced the concentration of astaxanthin by 1.3 times after 12 days in C. sorokiniana. It demonstrates that linoleic acid is a potent enhancer of astaxanthin production [125]. According to the study done by Yang ZY et al. [31], lipid production increased with the addition of phytohormones in saline wastewater cultivation of C. pyrenoidosa. The results showed an increase in lipid productivity by 1.8, 1.9, 2.5, 2.5, and 2.6 times by the application of brassinolide, gibberellins, indoleacetic acid, salicylic acid, and jasmonic acid, respectively. Application of indole propionic acid (10 mg/L) and indoleacetic acid (7.8 mg/L) increased the astaxanthin production and lipid content in Chromochloris zofingiensis [126]. Similarly, Scenedesmus obliquus showed 1.9 times increase in growth when applied with indole-3-acetic acid (IAA) (50 µM) [120]. A mixotrophic cultivation of C. vulgaris composed of 10 mg/L IAA, 2250 mg/L sodium nitrate and 10 g/L glucose under irradiation with red light resulted in high lipid content and biomass production [122]. ABA treatment upregulates fabF and fabI (fatty acid elongation), and desaturase genes like desA1 and desC, enhancing SFA and MUFA levels. Genes encoding antioxidant enzymes such as peroxiredoxin and RNA helicase DOB1 are also induced, helping combat oxidative stress [127]. Salicylic acid and high sodium acetate (SAHS) treatment significantly enhanced carotenoid and fatty acid accumulation in H. pluvialis, particularly during the early response phase. The upregulated genes included PSY, PDS, LCY, CRTO, CRTR-B, and BKT in the carotenoid biosynthesis pathway, and BC, ACP, MCTK, FAFA, FAD, and SAD in the fatty acid biosynthesis pathway [128]. In Chlorella sp. IAA treatment significantly upregulated acp, mctk, fata, and fad genes, with fad showing the highest expression, correlating with a marked increase in omega-3 fatty acid production and abscisic acid (ABA) also strongly induced acp, mctk, and fata gene expression, along with elevated ACC activity, enhancing total fatty acid synthesis [129].
Metabolic engineering in microalgae for lipid biosynthesis enhancement
Efforts have been directed towards enhancing photosynthetic efficiency to improve the growth rate. By constructing strains with improved photosynthetic efficiency, microalgae can potentially increase their biomass production and, consequently, lipid synthesis [53]. However, it is important to note that photosynthetic efficiency also directly impacts lipid synthesis. To further enhance lipid synthesis, specific strategies have been developed to improve the liposynthetic machinery [130]. One approach involves increasing the expression of ACC, which has been shown to stimulate lipid synthesis in microalgae [131]. By upregulating ACC expression, researchers aim to promote the conversion of acetyl-CoA to malonyl-CoA, a key precursor in lipid biosynthesis [132]. Some key genes involved in lipid biosynthesis, such as PLA2, ME2, DGAT2, ACC, and GPAT, and their targeted modifications to enhance lipid accumulation and metabolic flux regulation are illustrated in Fig. 5. Complementary to enhancing lipid synthesis, a strategy to repress lipid catabolism is also proposed. This can be achieved by downregulating or inhibiting processes such as TAG hydrolysis and β-oxidation, which are involved in the breakdown of lipids [133]. By limiting these pathways, microalgae can potentially redirect resources towards lipid synthesis, increasing lipid content in the cells.

Key genes and target sites involved in lipid biosynthesis in microalgae, highlighting their roles and potential genetic modifications to enhance production
In addition to manipulating lipid synthesis pathways, the regulation and insertion of specific desaturases, elongases, and associated fatty acid transporters (FATs) have been explored [134, 135]. These modifications can alter the fatty acid profile of microalgae, allowing for the production of desired lipid fractions with specific compositions [136]. Interestingly, these manipulations may also impact the overall lipid content or the biosynthesis of specific lipid fractions, offering a dual benefit in terms of lipid quality and quantity. The overexpression of the plastidial ACC gene (ACC1) in diatoms Cyclotella cryptica and Navicula saprophila did not result in improved fatty acid synthesis [137]. This suggests that upregulating ACC alone may not be sufficient to enhance lipid biosynthesis, indicating the need for a synergistic action of multiple enzymes. In light of the limited success in overexpressing genes involved in fatty acid synthesis, researchers shifted their attention to other enzymes implicated in acylglycerol biosynthesis, including both storage and structural forms. However, when DGAT genes (CrDGAT2a, CrDGAT2b, and CrDGAT2c) were overexpressed in the model microalga Chlamydomonas reinhardtii, increased lipid accumulation was not observed [138].
Interestingly, a study by Deng et al. [139] on five homologous CrDGAT2 genes in C. reinhardtii revealed diverse effects on lipid accumulation. Silencing of CrDGAT2-1 or CrDGAT2-5 led to a significant decrease in lipid content, while transformants harboring CrDGAT2-4 showed an increase in lipid content. No significant changes were observed when CrDGAT2-2 or CrDGAT2-3 was silenced. Moreover, in the diatom Phaeodactylum tricornutum, overexpression of a type 2 DGAT gene resulted in a 35% increase in TAG (triacylglycerol) accumulation [140]. This finding highlights the potential of manipulating specific DGAT genes to enhance lipid production in microalgae. Elongases and desaturases are primarily responsible for PUFA biosynthesis and not directly implicated in lipid accumulation, their regulation is a major target for altering unsaturation profiles in microalgae [141]. Characterization of elongase and desaturase genes in microalgae has provided valuable insights, although engineering trials are still in the early stages. Many efforts have focused on modifying the fatty acid composition, particularly to increase the content of omega-3 or omega-6 fatty acids.
However, most of these endeavors have primarily been conducted in transgenic plants, with limited progress in microalgae. Excitingly, a recent study by Hamilton et al. [142] showcased significant advancements in microalgal genetic engineering. They achieved an impressive eightfold increase in DHA content by expressing a heterologous Δ5 elongase from the Picoalga ostreococcus tauri in the diatom Phaeodactylum tricornutum. Additionally, overexpressing an annotated Δ5 desaturase gene (PtD5b) led to a remarkable 65% increase in lipid content and a 58% boost in EPA synthesis in P. tricornutum. Furthermore, studies have revealed the impact of specific desaturases on lipid fraction biosynthesis in microalgae. For instance, manipulating a Chlamydomonas gene encoding a Δ4 desaturase resulted in altered levels of monogalactosyldiacylglycerol (MGDG) [143]. Reduced enzyme levels led to a decrease in MGDG, while its overproduction increased the levels of 16:4 acyl groups in cell extracts and the total amount of MGDG. Apart from desaturases, FATs also play a role in shaping the fatty acid composition of microalgal lipids. Recent successes include the engineering of plant-derived FATs, such as the C12:0 and C14:0 specific FATs from Umbellularia californica and Cinnamomum camphora, respectively, into the diatom P. tricornutum [144]. This genetic modification effectively redirected fatty acid synthesis towards the biosynthesis of C12:0 and C14:0 fatty acids.
Astaxanthin: structure and enhancement strategies
Carotenoids are mostly 40-carbon terpenoids with eight isoprenoid units as their basic structural unit and responsible for yellow, orange, purple, and red colors in nature and are tetraterpenes widely distributed in photosynthetic organisms [145, 146] (Fig. 6). These pigments play key roles in photosynthesis by protecting cells from excessive light acting as antioxidants and contributing to photoprotection and structural components of light-harvesting complexes. Carotenoids are categorized into two classes: carotenes and xanthophylls. Carotenes, like β-carotene and lycopene, are hydrocarbon carotenoids, while xanthophylls such as lutein and zeaxanthin are oxygenated derivatives [147] (Fig. 7). Astaxanthin, a xanthophyll carotenoid is chemically characterized by its structure as 3,3′-dihydroxy-β, β′-carotene-4,4′-dione (CAS no. 472-61-7; molecular weight = 596.8 g/mol). It consists of a polyene chain connecting two β-ionone rings at its ends, each containing hydroxyl groups at the 3,3′ positions which create asymmetry. The biosynthesis of astaxanthin begins with the precursor isopentenyl pyrophosphate (IPP), derived from the methylerythritol 4-phosphate (MEP) pathway in prokaryotes and plastids [148] (Fig. 8). IPP and its isomer dimethylallyl pyrophosphate (DMAPP) serve as the fundamental building blocks for carotenoid biosynthesis. Through the action of prenyl transferases, geranylgeranyl pyrophosphate (GGPP) is synthesized by the sequential addition of IPP to farnesyl pyrophosphate (FPP) catalyzed by GGPP synthase [149]. Phytoene desaturase (PDS) and ζ-carotene desaturase (ZDS) then catalyze the conversion of colorless phytoene into lycopene [150]. The desaturation introduces conjugated double bonds, forming intermediate carotenoids such as ζ-carotene and neurosporene [151]. Lycopene is further converted into different carotenoids by the cyclization of its ends into ionone rings. The formation of β-carotene, which is the precursor of astaxanthin, involves two ε-cyclization reactions catalyzed by lycopene β-cyclase (LCY-b) [152]. In plants and certain algae, β-carotene acts as a crucial pigment for photosynthesis and a precursor for keto-carotenoids [153]. The biosynthesis of astaxanthin from β-carotene involves two enzymatic modifications: hydroxylation at the C3 position and ketolation at the C4 position on each of the β-ionone rings. A β-carotene hydroxylase introduces two hydroxyl groups, while a β-carotene ketolase adds two 4-keto groups, forming astaxanthin through these stepwise reactions.

Different types of carotenoids produced by different organisms

Two classes of carotenoids characterized on basis of cyclization and oxygen derivation

Biosynthetic pathway of astaxanthin production in microalgae DXP: 1-deoxy-D-xylulose-5-phosphate; PDS: phytoene desaturase; ZDS: zeta desaturase; LCY: lycopene cyclase; MEP: 2-C-methyl-D-erythritol-4-phosphate; IPP: isopentenyl pyrophosphate; DXS: 1-deoxy-D-xylulose-5-phosphate synthase; GGPP: geranyl geranyl diphosphate; DMAPP: Dimethyl-pyrophosphate; PSY: phytoene synthase
Strategies to enhance astaxanthin production in microalgae
Effect of light and nutrition
Two commonly employed cultivation modes for microalgae growth and astaxanthin production are autotrophic and mixotrophic approaches (Fig. 9). In photoautotrophic conditions, maintaining an appropriate CO2 concentration is crucial for Chlorella zofingiensis growth. Excessive CO2 levels can lower the pH of the culture medium, impeding algal growth, while insufficient CO2 concentrations fail to provide an adequate carbon source for effective photosynthesis [154]. Studies conducted in closed systems have determined that CO2 concentrations ranging from 1 to 10% result in optimal dry biomass productivity [154]. Additionally, nitrogen availability plays a pivotal role in the accumulation of astaxanthin, a secondary carotenoid, within autotrophic microalgae. Nitrogen starvation has been shown to enhance astaxanthin biosynthesis in C. zofingiensis [154].

Different strategies to enhance astaxanthin production
The bioproduction of astaxanthin typically involves two key steps: cell development and astaxanthin synthesis. Astaxanthin synthesis is induced under various stress conditions, including nutrient deprivation under specific light conditions, as well as manipulation of light intensities and colors, all of which significantly influence astaxanthin production [154]. To optimize astaxanthin accumulation, a two-phase strategy is commonly employed. Initially, low light intensities are utilized to facilitate microalgae growth until reaching the desired cell density. Subsequently, in the second phase, high light intensity is applied as a stressor to induce astaxanthin accumulation [155]. Among different light colors, blue wavelengths have been observed to elicit the highest astaxanthin accumulation compared to white and red light. This phenomenon can be attributed to photosystem II’s pigment–protein complexes receiving less light at blue wavelengths, minimizing photo-destruction potential and reducing photo-inhibition, thus promoting astaxanthin synthesis.
Cultivation systems and strategies
Photobioreactors are closed systems designed to optimize light and nutrient availability, providing better control over cultivation conditions. These systems offer advantages such as increased biomass productivity and improved astaxanthin synthesis. By regulating light intensity, photoperiod, and other environmental parameters, photobioreactors create favorable conditions for microalgae growth and astaxanthin accumulation. In a study by Le-Feuvre et al. [156], a tubular photobioreactor was used for astaxanthin production by H. pluvialis. The authors demonstrated that the controlled environment of the photobioreactor significantly enhanced the growth rate and astaxanthin content of the microalgae. The optimal cultivation conditions inside the photobioreactor, including light intensity, temperature, and CO2 concentration, resulted in a higher astaxanthin yield compared to open pond systems. Aeration also plays a crucial role in closed cultivation systems, as it can influence the growth rates of some microalgae. The turbulent flow and bubbles formed during aeration help to enhance gas–liquid contact and diffusion of gases, improving the uptake of nutrients [157]. However, high aeration rates can also induce hydrodynamic stress, leading to cell damage and death. The effects of aeration depend on several factors, such as algal strain, culture conditions, including pH, temperature, ionic strength, and salinity of the medium [158]. Studies have been conducted on the influence of aeration on microalgal biomass and lipid accumulation. A study conducted by Magdaong et al. [159] showed that the aeration rate and light cycle affect the growth of Chlorella sorokiniana in a photobioreactor. Results showed that the growth rate was highest for C. sorokiniana at 0.1000 vvm and continuous lightning [159]. A study by Goswami et al. [160] showed that aeration, along with optimized pH resulted in high biomass production and lipid biosynthesis in Tetraselmis indica [160]. Another study by Goswami, Mehariya et al. [162] on Picochlorum sp. BDUG 100241 (PC) demonstrated that a 12:12 h day–night cycle with pH of 7.0 along with aeration resulted in improved biomass production (2.66 g/L) and lipid content (44.04%) by showing highest nutrient removal efficiency with bioflocculation (Aspergillus niger MTCC 281 and Stropharia sp. ITCC-8422) approach. Mansouri et al. [158] demonstrated that aeration had no significant effect on biomass in Scenedesmus obliquus under photo tropical conditions, but an increase in lipid content was observed.
Mixotrophic cultivation is another cultivation strategy that has shown promise in improving astaxanthin yields. Mixotrophic cultivation provides organic carbon sources (heterotrophic mode) and light energy (autotrophic mode) to the microalgae. This approach allows for enhanced astaxanthin production by providing additional carbon substrates for metabolism, which can lead to increased biomass and astaxanthin synthesis. Oslan et al. [163] investigated the impact of mixotrophic cultivation on astaxanthin production in Chlorella zofingiensis. The study demonstrated that mixotrophic conditions, with the addition of glucose as a carbon source, resulted in higher astaxanthin content compared to autotrophic cultivation alone. Providing organic carbon stimulated the metabolic pathways involved in astaxanthin biosynthesis, leading to improved yields. By utilizing photobioreactors to optimize cultivation conditions and implementing mixotrophic strategies to enhance carbon availability, researchers have successfully increased astaxanthin production. These advancements offer better control over the cultivation environment and nutrient availability, ultimately leading to improved growth and astaxanthin synthesis in microalgae.
Nutrient optimization
Nutrient optimization plays a crucial role in enhancing astaxanthin production in microalgae. Manipulating the composition and concentration of growth media, particularly nitrogen and carbon sources, can significantly impact astaxanthin biosynthesis. Understanding the scientific reasons behind nutrient optimization is key to achieving higher yields. Nitrogen is an essential nutrient for microalgae growth and astaxanthin synthesis. Nitrogen availability can influence the metabolic pathways and regulatory mechanisms involved in astaxanthin biosynthesis. Nitrogen limitation or starvation has been found to induce astaxanthin accumulation in microalgae cells as a protective mechanism against stress [164]. Microalgae can shift their metabolic priorities towards astaxanthin synthesis by reducing nitrogen availability in the cultivation medium.
Nitrogen limitation triggers a series of molecular responses and metabolic alterations that redirect carbon resources towards secondary metabolite production, including astaxanthin [154]. For example, nitrogen starvation was applied to Chlorella zofingiensis cultures, enhancing astaxanthin biosynthesis [154]. Nitrogen-deprived conditions led to a significant increase in astaxanthin content, demonstrating the role of nitrogen limitation in promoting astaxanthin production. Carbon availability also influences astaxanthin biosynthesis in microalgae. Manipulating the carbon source in the growth medium can impact the carbon flux and allocation within the cell, affecting the synthesis of astaxanthin precursors. Carbon substrates, such as glucose or acetate, can be utilized as alternative carbon sources to enhance astaxanthin production in microalgae [165]. By providing additional carbon sources, microalgae can allocate more resources towards the synthesis of astaxanthin. This is because the availability of carbon substrates supports the energy and carbon requirements for astaxanthin biosynthesis pathways. The diversion of carbon resources towards astaxanthin synthesis under mixotrophic conditions has been shown to result in higher astaxanthin yields compared to autotrophic cultivation [165].
Nanoparticle
Nanoparticles have shown promising effects in advancing astaxanthin bioprocessing by enhancing microalgae growth, increasing astaxanthin production, and improving overall bioprocess efficiency [166,167,168]. The application of nanoparticles in astaxanthin bioprocessing involves various mechanisms and approaches, including nanoparticle-mediated delivery of nutrients, light manipulation, and stress alleviation. The application of silver nanoparticles (AgNPs) for enhanced astaxanthin production in microalgae has been investigated. Nasri et al. [169] studied the effect of different concentrations of AgNPs on astaxanthin biosynthesis in H. pluvialis. They found that the addition of AgNPs at a concentration of 10 ppm resulted in the highest carotenoid production. In their study, they initially observed a temporary growth inhibition effect of AgNPs. However, after acclimatization, the algal biomass increased, and the highest astaxanthin content was achieved at the 10 ppm concentration of AgNPs. This suggests that AgNPs can induce stress conditions that trigger astaxanthin biosynthesis in H. pluvialis.
The mechanism behind the enhanced astaxanthin production with AgNPs is not fully understood in this specific reference. However, nanoparticles like AgNPs have been reported to induce oxidative stress, activate stress response pathways, and modulate gene expression related to astaxanthin biosynthesis in microalgae [170]. It is important to note that the effects of nanoparticles on astaxanthin production can be influenced by various factors, such as nanoparticle size, concentration, surface properties, and specific microalgal species used in the study. Further research is needed to fully understand the underlying mechanisms and optimize the application of nanoparticles for astaxanthin bioprocessing [171].
Mutation
Mutation can play a crucial role in astaxanthin bioprocessing by introducing genetic variations that enhance astaxanthin production in microalgae. Through targeted mutations, specific genes involved in astaxanthin biosynthesis or related metabolic pathways can be modified, leading to improved astaxanthin yields. Several studies have utilized mutation techniques in astaxanthin bioprocessing, resulting in increased astaxanthin production. For example, Wang et al. [172] conducted a study where they overexpressed the crtR-B gene in H. pluvialis, a gene encoding a key enzyme in the astaxanthin biosynthesis pathway. By introducing a constitutive promoter to drive the expression of crtR-B, they observed a significant increase in astaxanthin content compared to the wild-type strain. Some of the key genes involved in carotenoid biosynthesis, such as PDS, bkt, dxs, psy, and ZEP, and their targeted modifications to enhance the production of astaxanthin, fucoxanthin, and β-carotene are illustrated in Fig. 10.

Key genes and target sites involved in lipid biosynthesis in microalgae, highlighting their roles and potential genetic modifications to enhance production
Xiong et al. [173] used CRISPR/Cas9 genome editing technology to mutate genes involved in astaxanthin biosynthesis in Chlorella zofingiensis. They targeted bkt1 and crtZ genes, which are critical for astaxanthin synthesis, resulting in a knockout mutant with reduced chlorophyll and increased astaxanthin accumulation compared to the wild-type strain. These examples illustrate how targeted mutations in genes associated with astaxanthin biosynthesis can improve astaxanthin production in microalgae (Table 2). However, it is important to carefully consider the potential impacts on the overall physiology and stability of the microalgae strains when performing genetic modifications. Additionally, optimizing the genetic modifications in conjunction with cultivation conditions and other process parameters is essential for maximizing astaxanthin bioproduction.
Relationship between fatty acid biogenesis and astaxanthin biosynthesis
Astaxanthin is synthesized from acetyl-CoA via IPP, phytoene, lycopene, and β-carotene, with the final step being the introduction of oxygen [174]. Phytoene saturase (PSY), phytoene desaturase, lycopene cyclase, β-carotene ketolase, and β-carotene hydroxylase (CrtR-b) are essential for astaxanthin production. Fatty acid biosynthesis in microalgae and plants initiates in the chloroplast and starts with the conversion of acetyl-CoA to malonyl-CoA [182]. These two pathways include characteristic enzymes like PSY, CrtR-b, biotin carboxylase, and stearoyl acyl carrier protein desaturase (SAD) in astaxanthin and fatty acid biosynthesis, respectively. In Haematococcus pluvialis, about 95% of astaxanthin molecules are esterified with fatty acids and stored in TAG-rich lipid bodies [183]. The synthesis of astaxanthin and fatty acids appears to be tightly coordinated, although the exact mechanisms behind this coordination are still unclear. Studies utilizing specific inhibitors such as norflurazon and cerulenin, which target β-carotene and fatty acid synthesis, respectively, demonstrate a feedback regulation between these pathways [184]. Inhibition of carotenogenesis causes a sharp decline in fatty acid biosynthesis, leading to the accumulation of excess fatty acids. Conversely, when fatty acid biosynthesis is suppressed, free astaxanthin inhibits further carotenogenesis. The esterification of astaxanthin relieves this feedback inhibition, allowing for its sequestration in cytoplasmic lipid bodies, facilitating coordination between the chloroplast and the endoplasmic reticulum. This coordination enhances carotenogenesis and fatty acid biosynthesis, especially under stress conditions, leading to increased astaxanthin accumulation. Also, TAG within lipid bodies acts as a solvent for astaxanthin esters, and the inhibition of TAG synthesis under high light conditions contributes to reduced astaxanthin production [128]. The location of the biosynthesis of astaxanthin is controversial. According to previous studies, β-carotene is synthesized in the chloroplast and exported to synthesize astaxanthin [185]. While it is also demonstrated that β-carotene synthesize free astaxanthin and its ester in association with endoplasmic reticulum [184]. A recent Raman microscopy of H. pluvialis reveals β-carotene synthesized in both the chloroplast and cytosolic lipid bodies, while astaxanthin is primarily concentrated in cytosolic lipid bodies [186]. In H. pluvialis ER-bound enzymes likely to use chloroplast-exported β-carotene for astaxanthin biosynthesis and esterification, as confirmed by immunoblot results and in vitro assays. In large mature LBs, the astaxanthin biosynthetic enzymes may continue to convert β-carotene to astaxanthin, which is subsequently esterified to astaxanthin esters [128]. Gao et al. [187] and others revealed that key genes involved in carotenoid biosynthesis, including PSY, PDS, ZDS, and CRTZ, are upregulated under stress conditions like salicylic acid and jasmonic acid inductions, causing increased astaxanthin accumulation. Transcriptional regulation of astaxanthin and lipid biosynthesis in H. pluvialis is analyzed under various stress conditions like salicylic acid, high light, and sodium acetate. Transcriptome-based analysis under these stress conditions identified key genes involved in both biosynthetic pathways, such as PSY, PDS, ZDS, BC, and ACP, that were upregulated in response to stress, leading to increased accumulation of astaxanthin and lipids [128]. Protein–protein interaction (PPI) analysis and Pearson correlation of outcome data revealed positive correlations among these genes. Positive correlation clearly suggests that there is a coordinated regulation at the transcriptional level [128].
Astaxanthin esterification may involve DGAT family enzymes, with DGAT1 and DGAT2 performing key acyltransferase functions. DGAT1 may transfer acyl groups from acyl CoA to retinol, producing retinyl esters from carotenoid degradation [188], and DGAT2 may use a fatty alcohol as CoA acceptor to produce a wax ester [189]. The DGAT1 inhibitor A922500 reduced astaxanthin synthesis by 55%, while xanthohumol fully blocked ester formation, causing free astaxanthin accumulation and hinting at minor contributions from other enzymes [128]. To further confirm the contribution of DGATs to astaxanthin esterification, molecular cloning of DGAT genes (including the DGAT1, DGAT2, PEP1 and unknown acyltransferases with DGAT activities) and characterization of the corresponding proteins are necessary. In general, microalgae have multiple DGAT homologs [190]. For instance, Chlamydomonas has one DGAT1 and five DGAT2 enzymes, and Nannochloropsis has two DGAT1 and 11 DGAT2 enzymes [190, 191]. DGATs transfer acyl groups to astaxanthin, linking carotenogenesis and fatty acid biosynthesis. β-Carotene to astaxanthin ester conversion also occurs in lipid bodies, where ER-resident enzymes are recruited.
Economic implications of integrating astaxanthin coproduction with biofuel manufacturing
Microalgae are one of the promising eco-friendly alternatives for the coproduction of astaxanthin and biofuel [192]. The biorefinery approach has become evident as an integrated method for utilizing microalga feedstock into high-value products including biofuels [193]. There is a requirement to develop an effective and profitable optimization process in order to obtain multiple complementary product production systems [194]. However, the per capita cost of microalgae biofuel production is not economically profitable [195]. An integrated biorefinery should be developed to enhance product outputs by using a single natural feedstock for the coproduction of multiple end products, which can make the process economically viable to some extent. Advanced technology has been implemented toward harvesting, cultivation, and enhancement of extraction method [196].
A two-stage cultivation approach has been applied for commercial production of astaxanthin from microalgae. Nutrients and a favorable environment are provided for high biomass production in the first stage, whereas in the second stage, stress conditions like nitrogen deficiency or high light are given for carotenoid induction, focusing on astaxanthin [197]. According to the study of Yu et al. [198], in a two-stage cultivation approach, low NaCl promotes the growth of H. pluvialis in the first stage and induces astaxanthin in high doses in the second stage.
Microalgal cultivation needs a high amount of fresh water and nutrients for large-scale production which is an issue for economical large-scale production. But microalgae can also utilize wastewater as a nutrient-rich source and this could serve the dual work, and produce valuable algal feedstock while removing nutrients from wastewater [161]. Wastewater is a nutrient-rich resource containing nitrogen and phosphorus, essential for microalgal growth, thereby lowering production costs and environmental impact. Species such as Haematococcus pluvialis and Chlorella zofingiensis are valued for their high astaxanthin content and accumulation of carbohydrates and lipids, which can be enhanced under stress conditions induced by wastewater components. Therefore, microalgal biomass cultivated in wastewater can be utilized to produce biogas, bioethanol, biodiesel, and fertilizer, among other products [199, 200]. Astaxanthin has uses in aquaculture feed, cosmetics, and nutraceuticals; nevertheless, contamination should be appropriately eliminated for humans to reduce safety hazards. Biodiesel is completely safe as wastewater contaminants do not affect biofuel composition but requires appropriate processing techniques to ensure the biofuel quality.
H. pluvialis cultivated in walnut shell extract showed improved lipid and astaxanthin production by 77.5 and 23.3%, respectively, along with wastewater recycling [198]. Culturing the microalgae in wastewater and aquafarming for photobioreactors, protein, astaxanthin, and exopolysaccharides leads to a reduction in the cost of biodiesel. Integrating astaxanthin production can lower biodiesel costs, reducing the prices from $3.90 to $0.54 per liter [201]. In the process of carotenogenesis, H. pluvialis also forms neutral lipids in the form of lipid droplets in the cytoplasm [202]. Therefore, the coproduction of lipid and astaxanthin could enable the H. pluvialis and C. zofingiensis feed stock for biodiesel production [203]. Most of the fatty acids in these species contain (16C to 18C) with the required saturation degree for biodiesel formation [204]. The biochemistry of biomass is affected by the growth conditions of the culture, so the selection of suitable stress as an inducer for lipid and astaxanthin accumulation is a deciding factor in microalgae biorefinery [205]. A significant loss of lipid and astaxanthin productivity was observed in H. pluvialis and C. zofingiensis during astaxanthin extraction, so to minimize product loss and contamination, a selective and separate extraction procedure for lipid and astaxanthin should be developed. Astaxanthin extraction using dimethyl ether (DME) and acetone was performed, and it was found that the DME extract consists of 0.33% astaxanthin and 1.82% in the acetone extract [206]. These results confirm that selective extraction can make a separate recovery of metabolite and lipid by the biorefinery approach. Research on common substrates such as pyruvate, which links the biosynthesis of both fatty acids and astaxanthin, offers another avenue for cost reduction [207]. Increasing CO2 levels or applying glycerol has been shown to boost both astaxanthin and lipid production [208].
Use of CRISPR/Cas 9 in metabolic engineering in microalgal system
The CRISPR/Cas9 system, particularly the widely utilized type II CRISPR/Cas9 system from Streptococcus pyogenes, stands as a cornerstone in the realm of genetic engineering. Originating from bacterial and archaeal adaptive immune systems, CRISPR/Cas systems offer unparalleled potential for precision genome editing. By leveraging short palindromic repeats interspaced with unique DNA fragments, these systems equip organisms with defense mechanisms against invasive nucleic acids like viruses, primarily achieved through the sequence-specific cleavage of foreign DNA [209]. This innovative immunity is established by incorporating short segments of invader DNA, known as spacers, within CRISPR loci. Upon encountering intruding DNA, these CRISPR arrays, containing the acquired spacers, undergo transcription, eventually leading to the formation of small interfering CRISPR RNAs (crRNAs) that are approximately 40 nucleotides long [210].
Teaming up with the transactivating CRISPR RNA (tracrRNA), these crRNAs take on the task of activating and guiding the Cas9 nuclease, a key player in the system. The dynamic interaction between crRNA and tracrRNA ignites the activity of Cas9, paving the way for precise genomic manipulation [211]. The prowess of CRISPR/Cas9 in cleaving homologous double-stranded DNA sequences, protospacers in invading DNA, is a groundbreaking feature. This molecular tool operates under the strict condition in which a protospacer-adjacent motif (PAM) is present downstream of the target DNA [212] (Fig. 11). In most cases, this PAM exhibits a distinct sequence, 5′-NGG-3′, which essentially serves as a molecular recognition tag enabling Cas9 to discern its designated cleavage site. The adaptability and simplicity of the CRISPR/Cas9 system distinguish it from previous genome editing techniques such as zinc finger nucleases (ZFNs) and transcription activator-like effector nucleases (TALENs), which necessitate intricate protein engineering. The CRISPR/Cas9 system’s reliance on a single-guide RNA (sgRNA) obviates the need for elaborate protein design, rendering the entire process more accessible and cost-effective [213]. Furthermore, this system’s potential for multiplex genome editing, achievable by employing multiple sgRNAs, presents a remarkable advantage compared to its predecessors.

General mechanism of genome editing by CRISPR–Cas9
Advantages of using CRISPR/Cas9 over ZFN’s and TALEN’s
The CRISPR/Cas9 system has emerged as a superior genome editing technique when compared to its predecessors, ZFNs and TALENs. This superiority can be attributed to CRISPR/Cas9’s remarkable precision, simplicity, and versatility, all of which are of immense significance in the realm of genetic engineering in microalgae. One of the standout advantages of CRISPR/Cas9 lies in its ability to achieve higher precision and accuracy in targeting specific DNA sequences [214]. This is achieved through the use of a short guide RNA (sgRNA) that guides the Cas9 nuclease to the desired DNA site via base pairing. This meticulous targeting minimizes the risk of off-target effects, where unintended modifications occur at unrelated sites in the genome. In contrast, ZFNs and TALENs rely on protein–DNA interactions, which can be more prone to off-target cleavage and pose challenges in maintaining accuracy [215]. An illustrative example of this precision in microalgae is evident in a study by Wang et al. [216], where CRISPR/Cas9 was used to target the fatty acid desaturase gene in Nannochloropsis oceanica. The study showcased high editing efficiency and the absence of significant off-target effects, leading to the creation of strains with enhanced lipid production. The simplicity and design flexibility of CRISPR/Cas9 further set it apart from its predecessors. The system’s reliance on RNA–DNA base pairing simplifies the design and construction process. Customizing sgRNA sequences to target specific genes is a straightforward procedure, allowing researchers to efficiently engineer the system [217].
In contrast, ZFNs and TALENs necessitate the complex and time-consuming design and assembly of custom DNA-binding proteins CRISPR/Cas9’s capacity for multiplex editing and high throughput represents another critical advantage. Its modular sgRNA design enables researchers to target multiple genes concurrently, a feature particularly useful for engineering complex metabolic pathways or studying gene interactions [218]. This multiplexing capability is achieved without the need for distinct protein designs for each target site, as is often the case with ZFNs and TALENs. Serif et al. [219] demonstrated this in microalgae by applying CRISPR/Cas9 to Phaeodactylum tricornutum to simultaneously knock out three genes involved in lipid metabolism. This streamlined approach exemplifies how CRISPR/Cas9 can accelerate metabolic engineering efforts. In addition to its technical advantages, CRISPR/Cas9 offers a cost-effective and accessible solution. The reliance on RNA-guided mechanisms reduces the necessity for labor-intensive protein engineering, resulting in overall cost savings [220]. This accessibility has led to widespread adoption of CRISPR/Cas9 across diverse laboratories and research settings.
In microalgae, Lin et al. [127] effectively harnessed CRISPR/Cas9 to enhance lipid content in Chlorella vulgaris by transforming the omega-3 fatty acid desaturase (fad3) gene. They highlighted the technology’s affordability and user-friendly nature, making it an attractive option for metabolic engineering studies. Another compelling advantage of the CRISPR/Cas9 system lies in its ability to significantly reduce the complexity of design and eliminate the intricate protein engineering required by ZFNs and TALENs. CRISPR/Cas9’s mechanism, driven by RNA–DNA base pairing, enables the straightforward customization of guide RNA sequences for precise gene targeting [221]. This streamlined approach contrasts with the demanding task of designing and assembling custom DNA-binding proteins for ZFNs and TALENs.
Strategies to overcome the technical bottlenecks that exist in microbial genetic engineering
The application of CRISPR/Cas9 for metabolic engineering in microalgae, particularly in enhancing lipid and astaxanthin biosynthesis, has both opportunities and challenges. One of the primary issues is the occurrence of off-target effects where the Cas9 endonuclease, guided by the sgRNA, cleaves unintended genomic loci. Microalgal genomes such as those of Amorphochlora amoeboformis and Lingulodinium polyedra species have a high degree of genetic redundancy and repetitive elements [222]. This genetic complexity increases the risk of off-target effects because the CRISPR/Cas9 system can mistakenly target homologous or similar sequences. The presence of paralogous genes further complicates this issue as the sgRNA may bind to multiple loci with high sequence similarity, leading to unintended double-strand breaks (DSBs) at these off-target sites [223]. This results in the disruption of non-target genes, which may have critical roles in cellular functions or metabolic pathways. Specificity of CRISPR/Cas9 is also influenced by the local genomic context, particularly the GC content of the target sequence. High GC content can enhance the binding affinity of sgRNAs not only at the intended site, but also at off-target loci with similar GC-rich sequences [224]. This increased affinity raises the likelihood of off-target cleavage events, which can introduce unintended mutations that disrupt gene function or regulatory elements. Also, epigenetic modifications such as DNA methylation, which are prevalent in microalgae, can alter the accessibility of genomic regions to the CRISPR/Cas9 machinery [225].
To address these challenges, several strategies have been developed to improve the specificity of CRISPR/Cas9 and reduce off-target effects. One of the most effective approaches is the design and optimization of sgRNAs. Advanced bioinformatics tools, such as CRISPR, CCTop, and CRISPRseek, enable the prediction and ranking of sgRNAs based on their specificity and the likelihood of off-target binding [226]. Detecting and mitigating off-target effects is essential in CRISPR/Cas9. Cas-OFFinder is a widely used tool that efficiently scans entire genomes for potential off-target sites by considering mismatches and bulges between the sgRNA and DNA target sequence [227]. CasAnalyzer (Cas01) integrates multiple prediction models, providing detailed reports and an optimized scoring system to rank the likelihood of off-target cleavage [228]. For high-throughput screening, CRISPR-Flash offers rapid identification of off-target sites across large datasets, leveraging a fast algorithm that efficiently predicts off-target effects based on sequence homology [229]. In the context of paired nickase applications, CROP-IT CFD calculates cutting frequency determination (CFD) scores to optimize sgRNA pair selection, offering a more accurate prediction of off-target activity [230]. DeepCRISPR advances the field further by employing deep learning models to provide probabilistic assessments of off-target sites, integrating sequence features and experimental data for high accuracy [231]. CRISP-Flash (Flashfry) is optimized for speed and scalability, making it ideal for large-scale CRISPR screens as it rapidly identifies and ranks potential off-target sites using a sophisticated scoring system. sgRNA design strategies that incorporate mismatches near the 5′ end of the guide sequence and a higher GC content near the 3′ end (PAM-proximal region) can enhance target specificity and reduce off-target binding [232]4.
Engineered Cas9 variants also offer a promising solution to the problem of off-target effects. Variants such as SpCas9-HF1 and eSpCas9(1.1) have been developed with mutations that reduce the non-specific interactions between the sgRNA and off-target DNA sequences [233, 234]. These engineered proteins exhibit reduced tolerance for mismatches between the sgRNA and the target DNA. The use of highly efficient CRISPR/Cas9, like SpCas9-HF1, increases precise monoallelic and biallelic editing events in primordial germ cell variants and significantly reduces off-target activity [235].
Another approach to minimizing off-target effects involves the use of paired Cas9 nickases. Unlike the standard Cas9, which induces DSBs, nickases introduce single-strand breaks on opposite DNA strands [236]. This dual nick approach requires two sgRNAs to bind adjacent sites on the target DNA, reducing the likelihood of off-target effects, as both nickases must simultaneously bind to their respective target sites. SpCas9-HF1 enhances the accuracy of cell cycle-dependent genome editing by increasing HDR efficiency and reducing off-target effects and indel rates [237]. Base editing technologies represent an additional strategy to reduce off-target effects. Base editors, which consist of a catalytically impaired Cas9 fused to a cytidine or adenosine deaminase, enable the precise conversion of single nucleotides without creating DSBs [238]. This approach circumvents the need for DNA repair processes that can introduce errors, thereby reducing the risk of off-target mutations. In microalgae, base editors have the potential to precisely modify key nucleotides in genes involved in lipid and astaxanthin biosynthesis, enhancing metabolic efficiency while minimizing unintended genetic alterations.
Comparative analysis of strategies for enhancing lipid and astaxanthin production
Various strategies have been developed to enhance lipid and astaxanthin production in microalgae, including environmental stress induction, nutrient modulation, metabolic engineering and cultivation system optimization. While environmental stress-based approaches such as salinity, high light intensity, and nutrient deprivation often stimulate lipid and astaxanthin accumulation by triggering oxidative stress and redirecting carbon flux towards storage compounds, they frequently compromise biomass productivity [239, 240]. In contrast, metabolic engineering allows precise targeting of key biosynthetic genes, such as DGAT for triacylglycerol synthesis and BKT or PSY for astaxanthin biosynthesis, thereby offering higher product yields with reduced growth penalties [219]. However, these methods demand advanced molecular tools and are subject to regulatory scrutiny. Nutrient modulation offers a moderate yet scalable solution, often used in tandem with cultivation optimization strategies like mixotrophy or photobioreactor systems to balance growth and product synthesis [61, 241]. Notably, the physiological overlap between lipid and astaxanthin biosynthesis, such as their shared precursors (e.g., acetyl-CoA) and common induction signals (e.g., ROS)—presents opportunities for synergistic enhancement.
Conclusion and future prospective
Microalgae, as unicellular photosynthetic organisms have become a viable option for the generation of biofuel in the renewable energy industry. Also, microalgae are a great source of various nutraceuticals and pharma products which have immense application for humankind purposes. This paper provided a thorough explanation of the many stress-induced tactics and genetic engineering methods used to increase the synthesis of lipids and astaxanthin. By combining stressors with cutting-edge molecular tools like CRISPR–Cas9, the potential for increasing yields and streamlining production routes has been greatly increased. Accepting these advances not only advances the biotechnological production of lipids and astaxanthin, but it also highlights the enormous potential of synthetic biology to transform the direction of sustainable bioresource production in the future. The synthesis of lipids and astaxanthin, is about to undergo a revolution thanks to the combination of genetic engineering and stress-induced techniques. Lipid-derived biofuels, with their optimized production processes and improved yields, provide a viable solution to the growing worldwide need for sustainable energy sources. In a similar vein, astaxanthin is becoming increasingly important in the pharmaceutical and nutraceuticals industries due to growing interest in its numerous health advantages. Future research could strategically integrate these developments to create integrated bioprocesses, utilizing the knowledge gained from this review to do so. This would boost the production of astaxanthin and biofuels while also sparking a new direction to a more sustainable and healthy future.
Data availability
No datasets were generated or analysed during the current study.
Abbreviations
- CO2 :
-
Carbon dioxide
- CRISPR:
-
Clustered regularly interspaced short palindromic repeats
- ROS:
-
Reactive oxygen species
- TAGs:
-
Triacylglycerols
- SQDG:
-
Sulfoquinovosyldiacylglycerol
- MGDG:
-
Monogalactosyldiacylglycerol
- PC:
-
Phosphatidylcholine
- PE:
-
Phosphatidylethanolamine
- G3P:
-
Glycerate-3-phosphate
- PDC:
-
Pyruvate dehydrogenase complex
- ACCase:
-
Acetyl-CoA carboxylase
- ACP:
-
Acyl carrier protein
- FAS:
-
Fatty acid synthase
- FAs:
-
Fatty acids
- SFAs:
-
Saturated fatty acids
- MUFAs:
-
Monounsaturated fatty acids
- PUFAs:
-
Polyunsaturated fatty acids
- FAS:
-
Fatty acid synthase
- FAT:
-
Fatty-ACP thioesterases
- PKS:
-
Polyketide synthase
- GPAT:
-
Glycerol-3-phosphate acyltransferase
- PA:
-
Phosphatidate
- LPA:
-
Lysophosphatidate
- EPA:
-
Eicosapentaenoic acid
- DHA:
-
Docosahexaenoic acid
- CCMs:
-
Carbon concentrating mechanisms
- P:
-
Phosphorus
- N:
-
Nitrogen
- Cu:
-
Copper
- Fe:
-
Iron
- Mn:
-
Manganese
- Ni:
-
Nickel
- Zn:
-
Zinc
- Cd:
-
Cadmium
- Cr:
-
Chromium
- Pb:
-
Lead
- Hg:
-
Mercury
- As:
-
Arsenic
- NaCl:
-
Sodium chloride
- MGDG:
-
Monogalactosyldiacylglycerol
- MEP:
-
Methylerythritol 4-phosphate
- DMAPP:
-
Dimethylallyl pyrophosphate
- GGPP:
-
Geranylgeranyl pyrophosphate
- FPP:
-
Farnesyl pyrophosphate
- PDS:
-
Phytoene desaturase
- ZDS:
-
ζ-Carotene desaturase
- AgNPs:
-
Silver nanoparticles
- PSY:
-
Phytoene saturase
- SAD:
-
Stearoyl acyl desaturase
- DME:
-
Dimethyl ether
- PAM:
-
Protospacer-adjacent motif
- ZFNs:
-
Zinc finger nucleases
- TALENs:
-
Transcription activator-like effector nucleases
- DSBs:
-
Double-strand breaks
- CFD:
-
Cutting frequency determination
References
-
Takács-György K, Lászlók A, Takács I. Examination of first generation biofuel production in some selected biofuel producing countries in Europe: a case study. Agric Econ Czech. 2020;66:469–76. https://doi.org/10.17221/237/2020-AGRICECON.
-
Sajedinia E, Tyner W. Use of general equilibrium models in evaluating biofuels policies. In: Dixon P, Francois J, van der Mensbrugghe D, editors. Policy analysis and modeling of the global economy. World Scientific Publishing; 2021. p. 437–65. https://doi.org/10.1142/9789811233630_0014.
-
Yin Z, Zhu L, Li S, Hu T, Chu R, Mo F, Hu D, Liu C, Li B. A comprehensive review on cultivation and harvesting of microalgae for biodiesel production: environmental pollution control and future directions. Bioresour Technol. 2020;301:122804. https://doi.org/10.1016/j.biortech.2020.122804.
-
Chowdury K, Nahar N, Deb U. The growth factors involved in microalgae cultivation for biofuel production: a review. Comput Water Eng Environ Eng. 2020;9:185–215. https://doi.org/10.4236/cweee.2020.94012.
-
Alalwan HA, Alminshid AH, Aljaafari HA. Promising evolution of biofuel generations. Subject review. Rene Ener Foc. 2019;28:127–39. https://doi.org/10.1016/J.REF.2018.12.006.
-
Chittora D, Meena M, Barupal T, Swapnil P, Sharma K. Cyanobacteria as a source of biofertilizers for sustainable agriculture. Biochem Biophys Rep. 2020;22:100737. https://doi.org/10.1016/j.bbrep.2020.100737.
-
Shin Y, Choi H, Choi J, Lee J, Sung Y, Sim S. Multilateral approach on enhancing economic viability of lipid production from microalgae: a review. Bioresour Technol. 2018;258:335–44. https://doi.org/10.1016/j.biortech.2018.03.002.
-
Hossain N, Zaini J, Mahlia T. Experimental investigation of energy properties for Stigonematales sp. microalgae as potential biofuel feedstock. Int J Sustain Eng. 2018;12(2):123–30. https://doi.org/10.1080/19397038.2018.1521882.
-
Murta A, Freitas M, Ferreira C, Peixoto M. The use of palm oil biodiesel blends in locomotives: an economic, social and environmental analysis. Renew Energy. 2021;164:521–30. https://doi.org/10.1016/J.RENENE.2020.08.094.
-
Rizwan M, Mujtaba G, Memon S, Lee K, Rashid N. Exploring the potential of microalgae for new biotechnology applications and beyond: a review. Renew Sust Energ Rev. 2018;92:394–404. https://doi.org/10.1016/J.RSER.2018.04.034.
-
Fernandes T, Cordeiro N. Microalgae as sustainable biofactories to produce high-value lipids: biodiversity, exploitation, and biotechnological applications. Mar Drugs. 2021;19(10):573. https://doi.org/10.3390/md19100573.
-
Santiago-Morales I, Trujillo-Valle L, Márquez-Rocha F, Hernández J. Tocopherols, phycocyanin and superoxide dismutase from microalgae: as potential food antioxidants. Appl Food Biotechnol. 2018;5:19–27. https://doi.org/10.22037/AFB.V5I1.17884.
-
González-Delgado AD, Barajas C, Morales J, Villabona R. Technical-economic prefeasibility study of astaxanthin production system from H. pluvial microalgae in Colombia. Indian J Sci Technol. 2018;11(34):1–8. https://doi.org/10.17485/IJST/2018/V11I34/122627.
-
Lai TT, Yang CM, Yang CH. Astaxanthin protects retinal photoreceptor cells against high glucose-induced oxidative stress by induction of antioxidant enzymes via the PI3K/Akt/Nrf2 pathway. Antioxidants. 2020;9(8):729. https://doi.org/10.3390/antiox9080729.
-
Gupta AK, Seth K, Maheshwari K, Baroliya PK, Meena M, Kumar A, Vinayak V, Harish. Biosynthesis and extraction of high-value carotenoid pigments from algae. Front Biosci Landmark. 2021;26(6):171–90. https://doi.org/10.52586/4932.
-
Zhang Y, Ye Y, Ding W, Mao X, Li Y, Gerken H, Liu J. Astaxanthin is ketolated from zeaxanthin independent of fatty acid synthesis in Chromochloris zofingiensis. Plant Physiol. 2020;183(3):883–97. https://doi.org/10.1104/pp.20.00325.
-
Ranjbar S, Malcata FX. Challenges and prospects for sustainable microalga-based oil: a comprehensive review, with a focus on metabolic and genetic engineering. Fuel. 2022;324:124567. https://doi.org/10.1016/j.fuel.2022.124567.
-
Ekpe L, Inaku K, Ekpe V. Antioxidant effects of astaxanthin in various diseases—a review. J Mol Pathophysiol. 2018;7(1):1–6. https://doi.org/10.5455/oams.20180315075538.
-
Mei S, Song X, Wang Y, Wang J, Su S, Zhu J, Geng Y. Studies on protection of astaxanthin from oxidative damage induced by H2O2 in RAW264.7 cells based on 1H NMR metabolomics. J Agric Food Chem. 2019;67(49):13568–76. https://doi.org/10.1021/acs.jafc.9b04587.
-
Sorrenti V, Davinelli S, Scapagnini G, Willcox B, Allsopp R, Willcox D. Astaxanthin as a putative Geroprotector: molecular basis and focus on brain aging. Mar Drugs. 2020;18(7):351. https://doi.org/10.3390/md18070351.
-
Yiğit M, Güneş A, Uğuz C, Yalçın T, Tök L, Öz A, Nazıroğlu M. Effects of astaxanthin on antioxidant parameters in ARPE-19 cells on oxidative stress model. Int J Ophthalmol. 2019;12(6):930–5. https://doi.org/10.18240/ijo.2019.06.08.
-
Zaafan M, Abdelhamid A. The cardioprotective effect of astaxanthin against isoprenaline-induced myocardial injury in rats: involvement of TLR4/NF-κB signaling pathway. Eur Rev Med Pharmacol Sci. 2021;25(11):4099–105. https://doi.org/10.26355/eurrev_202106_26052.
-
Fassett RG, Coombes JS. Astaxanthin, oxidative stress, inflammation and cardiovascular disease. Future Cardiol. 2009;5(4):333–42. https://doi.org/10.2217/FCA.09.19.
-
Wu D, Xu H, Chen J, Zhang L. Effects of astaxanthin supplementation on oxidative stress. Int J Vitam Nutr Res. 2019;90(1–2):179–94. https://doi.org/10.1024/300-9831/a000497.
-
Chernova N, Korobkova T, Kiseleva S. Use of biomass for producing liquid fuel: current state and innovations. Therm Eng. 2010;57:937–45. https://doi.org/10.1134/S0040601510110054.
-
Udayan A, Pandey AK, Sirohi R, Sreekumar N, Sang BI, Sim SJ, Pandey A. Production of microalgae with high lipid content and their potential as sources of nutraceuticals. Phytochem Rev. 2023;22(4):833–60. https://doi.org/10.1007/s11101-021-09784-y.
-
Zhu LD, Li ZH, Hiltunen E. Strategies for lipid production improvement in microalgae as a biodiesel feedstock. BioMed Res Int. 2016;2016:8792548. https://doi.org/10.1155/2016/8792548.
-
Gao P, Guo L, Zhao Y, Jin C, She Z, Gao M. Enhancing microalgae growth and product accumulation with carbon source regulation: new perspective for the coordination between photosynthesis and aerobic respiration. Chemosphere. 2021;278:130435. https://doi.org/10.1016/j.chemosphere.2021.130435.
-
Mirizadeh S, Nosrati M, Shojaosadati SA. Synergistic effect of nutrient and salt stress on lipid productivity of Chlorella vulgaris through two-stage cultivation. Bioenerg Res. 2020;13:507–17. https://doi.org/10.1007/s12155-019-10077-8.
-
Nymark M, Sharma A, Sparstad T, Bones A, Winge P. A CRISPR/Cas9 system adapted for gene editing in marine algae. Sci Rep. 2016;6:24951. https://doi.org/10.1038/srep24951.
-
Yang ZY, Huang KX, Zhang YR, Yang L, Zhou JL, Yang Q, Gao F. Efficient microalgal lipid production driven by salt stress and phytohormones synergistically. Bioresour Technol. 2023;367:128270. https://doi.org/10.1016/j.biortech.2022.128270.
-
Zhao Y, Wang Q, Gu D, Huang F, Liu J, Yu L, Yu X. Melatonin, a phytohormone for enhancing the accumulation of high-value metabolites and stress tolerance in microalgae: applications, mechanisms, and challenges. Bioresour Technol. 2023;393:130093. https://doi.org/10.1016/j.biortech.2023.130093.
-
Hu Q, Huang D, Li A, Hu Z, Gao Z, Yang Y, Wang C. Transcriptome-based analysis of the effects of salicylic acid and high light on lipid and astaxanthin accumulation in Haematococcus pluvialis. Biotechnol Biofuels. 2021;14(1):82. https://doi.org/10.1186/s13068-021-01933-x.
-
Hamed I. The evolution and versatility of microalgal biotechnology: a review. Compr Rev Food Sci Food Saf. 2016;15(6):1104–23. https://doi.org/10.1111/1541-4337.12227.
-
Young D, Shachar-Hill Y. Large fluxes of fatty acids from membranes to triacylglycerol and back during N-deprivation and recovery in Chlamydomonas. Plant Physiol. 2020;185(3):796–814. https://doi.org/10.1093/plphys/kiaa071.
-
Gabruk M, Myśliwa-Kurdziel B, Kruk J. MGDG, PG and SQDG regulate the activity of light-dependent protochlorophyllide oxidoreductase. Biochem J. 2017;474(7):1307–20. https://doi.org/10.1042/BCJ20170047.
-
Sengmee D, Cheirsilp B, Suksaroge T, Prasertsan P. Biophotolysis-based hydrogen and lipid production by oleaginous microalgae using crude glycerol as exogenous carbon source. Int J Hydrog Energy. 2017;42:1970–6. https://doi.org/10.1016/J.IJHYDENE.2016.10.089.
-
Dawaliby R, Trubbia C, Delporte C, Noyon C, Ruysschaert J, Antwerpen P, Govaerts C. Phosphatidylethanolamine is a key regulator of membrane fluidity in eukaryotic cells. J Bio Chem. 2015;291:3658–67. https://doi.org/10.1074/jbc.M115.706523.
-
Guarnieri MT, Nag A, Smolinski SL, Darzins A, Seibert M, Pienkos PT. Examination of triacylglycerol biosynthetic pathways via de novo transcriptomic and proteomic analyses in an unsequenced microalga. PLoS ONE. 2011;6(10):e25851. https://doi.org/10.1371/journal.pone.0025851.
-
Lee H, Son J, Sim S, Woo H. Metabolic rewiring of synthetic pyruvate dehydrogenase bypasses for acetone production in cyanobacteria. Plant Biotechnol J. 2020;18:1860–8. https://doi.org/10.1111/pbi.13342.
-
Danesh A, Mooij P, Ebrahimi S, Kleerebezem R, Loosdrecht M. Effective role of medium supplementation in microalgal lipid accumulation. Biotechnol Bioeng. 2018;115:1152–60. https://doi.org/10.1002/bit.26548.
-
Wang Q, Feng Y, Lu Y, Xin Y, Shen C, Wei L, Liu Y, Lv N, Du X, Zhu W, Jeong B, Xue S, Xu J. Manipulating fatty-acid profile at unit chain-length resolution in the model industrial oleaginous microalgae Nannochloropsis. Metab Eng. 2021;66:157–66. https://doi.org/10.1016/j.ymben.2021.03.015.
-
Wang Q, Lu Y, Xin Y, Wei L, Huang S, Xu J. Genome editing of model oleaginous microalgae Nannochloropsis spp. by CRISPR/Cas9. Plant J. 2016;88(6):1071–81. https://doi.org/10.1111/tpj.13307.
-
Milke L, Marienhagen J. Engineering intracellular malonyl-CoA availability in microbial hosts and its impact on polyketide and fatty acid synthesis. App Microbiol Biotechnol. 2020;104:6057–65. https://doi.org/10.1007/s00253-020-10643-7.
-
Gong Y, Huang J. Characterization of four untapped microalgae for the production of lipids and carotenoids. Algal Res. 2020;49:101897. https://doi.org/10.1016/j.algal.2020.101897.
-
Chen H, Wang Q. Regulatory mechanisms of lipid biosynthesis in microalgae. Biol Rev Camb Philos Soc. 2021;96(5):2373–91. https://doi.org/10.1111/brv.12759.
-
Xu Y. Biochemistry and biotechnology of lipid accumulation in the microalga Nannochloropsis oceanica. J Agric Food Chem. 2022;70(37):11500–9. https://doi.org/10.1021/acs.jafc.2c05309.
-
Südfeld C, Kiyani A, Wefelmeier K, Wijffels RH, Barbosa MJ, D’Adamo S. Expression of glycerol-3-phosphate acyltransferase increases non-polar lipid accumulation in Nannochloropsis oceanica. Microb Cell Fact. 2023;22(1):12. https://doi.org/10.1186/s12934-022-01987-y.
-
Song Y, Wang F, Chen L, Zhang W. Engineering fatty acid biosynthesis in microalgae: recent progress and perspectives. Mar Drugs. 2024;22(5):216. https://doi.org/10.3390/md22050216.
-
Blasio M, Balzano S. Fatty acids derivatives from eukaryotic microalgae, pathways and potential applications. Front Microbiol. 2021;12:718933. https://doi.org/10.3389/fmicb.2021.718933.
-
Huerlimann R, Heimann K. Comprehensive guide to acetyl-carboxylases in algae. Crit Rev Biotechnol. 2013;33(1):49–65. https://doi.org/10.3109/07388551.2012.668671.
-
Fan J, Andre C, Xu C. A chloroplast pathway for the de novo biosynthesis of triacylglycerol in Chlamydomonas reinhardtii. FEBS Lett. 2011;585(12):1985–91. https://doi.org/10.1016/j.febslet.2011.05.018.
-
Alishah Aratboni H, Rafiei N, Garcia-Granados R, Alemzadeh A, Morones-Ramírez JR. Biomass and lipid induction strategies in microalgae for biofuel production and other applications. Microb Cell Fact. 2019;18:1–17. https://doi.org/10.1186/s12934-019-1228-4.
-
Nisar S, Hanif MA, Rashid U, Hanif A, Akhtar MN, Ngamcharussrivichai C. Trends in widely used catalysts for fatty acid methyl esters (Fame) production: a review. Catalysts. 2021;11(9):1085. https://doi.org/10.3390/catal11091085.
-
Singh R, Arora A, Singh V. Biodiesel from oil produced in vegetative tissues of biomass—a review. Bioresour Technol. 2021;326:124772. https://doi.org/10.1016/j.biortech.2021.124772.
-
Stansell GR, Gray VM, Sym SD. Microalgal fatty acid composition: implications for biodiesel quality. J Appl Phycol. 2012;24:791–801. https://doi.org/10.1007/s10811-011-9696-x.
-
Pradana YS, Makertihartha IGB, Indarto A, Prakoso T, Soerawidjaja TH. A review of biodiesel cold flow properties and its improvement methods: towards sustainable biodiesel application. Energies. 2024;17(18):4543. https://doi.org/10.3390/en17184543.
-
Bucy HB, Baumgardner ME, Marchese AJ. Chemical and physical properties of algal methyl ester biodiesel containing varying levels of methyl eicosapentaenoate and methyl docosahexaenoate. Algal Res. 2012;1(1):57–69. https://doi.org/10.1016/j.algal.2012.02.001.
-
Udayan A, Sirohi R, Sreekumar N, Sang BI, Sim SJ. Mass cultivation and harvesting of microalgal biomass: current trends and future perspectives. Bioresour Technol. 2022;344:126406. https://doi.org/10.1016/j.biortech.2021.126406.
-
Shokravi Z, Shokravi H, Chyuan OH, Lau WJ, Koloor SSR, Petrů M, Ismail AF. Improving ‘lipid productivity’ in microalgae by bilateral enhancement of biomass and lipid contents: a review. Sustainability. 2020;12(21):9083. https://doi.org/10.3390/su12219083.
-
Zhang Y, Kang X, Zhen F, Wang Z, Kong X, Sun Y. Assessment of enzyme addition strategies on the enhancement of lipid yield from microalgae. Biochem Eng J. 2022;177:108198. https://doi.org/10.1016/j.bej.2021.108198.
-
Menegol T, Diprat AB, Rodrigues E, Rech R. Effect of temperature and nitrogen concentration on biomass composition of Heterochlorella luteoviridis. Food Sci Technol. 2017;37(spe):28–37. https://doi.org/10.1590/1678-457X.13417
-
Huang X, Huang Z, Wen W, Yan J. Effects of nitrogen supplementation of the culture medium on the growth, total lipid content and fatty acid profiles of three microalgae (Tetraselmis subcordiformis, Nannochloropsis oculata and Pavlova viridis). J Appl Phycol. 2013;25:129–37. https://doi.org/10.1007/s10811-012-9846-9.
-
Ota S, Oshima K, Yamazaki T, Kim S, Yu Z, Yoshihara M, Kawano S. Highly efficient lipid production in the green alga Parachlorella kessleri: draft genome and transcriptome endorsed by whole-cell 3D ultrastructure. Biotechnol Biofuels. 2016;9:13. https://doi.org/10.1186/s13068-016-0424-2.
-
Zhao Y, Li D, Ding K, Che R, Xu JW, Zhao P, Yu X. Production of biomass and lipids by the oleaginous microalgae Monoraphidium sp. QLY-1 through heterotrophic cultivation and photo-chemical modulator induction. Bioresour Technol. 2016;211:669–76. https://doi.org/10.1016/j.biortech.2016.03.160.
-
Abdelkarim OH, Wijffels RH, Barbosa MJ. Microalgal lipid production: a comparative analysis of Nannochloropsis and Microchloropsis strains. J Appl Phycol. 2024;37:15–34. https://doi.org/10.1007/s10811-024-03318-7.
-
Benasla A, Hausler R. A two-step cultivation strategy for high biomass production and lipid accumulation of Raphidocelis subcapitata immobilized in alginate gel. Biomass. 2021;1(2):94–104. https://doi.org/10.3390/biomass1020007.
-
Maltsev Y, Maltseva K, Kulikovskiy M, Maltseva S. Influence of light conditions on microalgae growth and content of lipids, carotenoids, and fatty acid composition. Biology. 2021;10(10):1060. https://doi.org/10.3390/biology10101060.
-
Sun H, Wu T, Chen SHY, Ren Y, Yang S, Huang J, Chen F. Powerful tools for productivity improvements in microalgal production. Renew Sust Energ Rev. 2021;152:111609. https://doi.org/10.1016/j.rser.2021.111609.
-
Pal D, Khozin-Goldberg I, Cohen Z, Boussiba S. The effect of light, salinity, and nitrogen availability on lipid production by Nannochloropsis sp. Appl Microbiol Biotechnol. 2011;90:1429–41. https://doi.org/10.1007/s00253-011-3170-1.
-
Pereira S, Otero A. Haematococcus pluvialis bioprocess optimization: effect of light quality, temperature and irradiance on growth, pigment content and photosynthetic response. Algal Res. 2020;51:102027. https://doi.org/10.1016/j.algal.2020.102027.
-
Zhong Y, Jin P, Cheng JJ. A comprehensive comparable study of the physiological properties of four microalgal species under different light wavelength conditions. Planta. 2018;248:489–98. https://doi.org/10.1007/s00425-018-2899-5.
-
Cha TS, Yee W, Phua PSP, Loh SH, Aziz A. A brief period of darkness induces changes in fatty acid biosynthesis towards accumulation of saturated fatty acids in Chlorella vulgaris UMT-M1 at stationary growth phase. Biotechnol Lett. 2021;43:803–12. https://doi.org/10.1007/s10529-021-03077-2.
-
Cheirsilp B, Torpee S. Enhanced growth and lipid production of microalgae under mixotrophic culture condition: effect of light intensity, glucose concentration and fed-batch cultivation. Bioresour Technol. 2012;110:510–6. https://doi.org/10.1016/j.biortech.2012.01.125.
-
Nzayisenga JC, Farge X, Groll SL, Sellstedt A. Effects of light intensity on growth and lipid production in microalgae grown in wastewater. Biotechnol Biofuels. 2020;13(4):4. https://doi.org/10.1186/s13068-019-1646-x.
-
Hosseini NS, Shang H, Scott JA. Biosequestration of industrial off-gas CO2 for enhanced lipid productivity in open microalgae cultivation systems. Renew Sust Energ Rev. 2018;92:458–69. https://doi.org/10.1016/j.rser.2018.04.086.
-
Makareviciene V, Andrulevičiūtė V, Skorupskaitė V, Kasperovičienė J. Cultivation of microalgae Chlorella sp. and Scenedesmus sp. as a potential biofuel feedstock. Environ Res Eng Manag. 2011;57(3):21–7. https://doi.org/10.5755/j01.erem.57.3.476.
-
Cheng J, Zhu Y, Zhang Z, Yang W. Modification and improvement of microalgae strains for strengthening CO2 fixation from coal-fired flue gas in power plants. Bioresour Technol. 2019;291:121850. https://doi.org/10.1016/j.biortech.2019.121850.
-
Mondal M, Khanra S, Tiwari ON, Gayen K, Halder GN. Role of carbonic anhydrase on the way to biological carbon capture through microalgae—a mini reviews. Environ Prog Sustain Energy. 2016;35(6):1605–15. https://doi.org/10.1002/ep.12394.
-
Mus F, Toussaint JP, Cooksey KE, Fields MW, Gerlach R, Peyton BM, Carlson RP. Physiological and molecular analysis of carbon source supplementation and pH stress-induced lipid accumulation in the marine diatom Phaeodactylum tricornutum. Appl Microbiol Biotechnol. 2013;97(8):3625–42. https://doi.org/10.1007/s00253-013-4747-7.
-
Barati B, Zeng K, Baeyens J, Wang S, Addy M, Gan SY, Abomohra AEF. Recent progress in genetically modified microalgae for enhanced carbon dioxide sequestration. Biomass Bioenergy. 2021;145:105927. https://doi.org/10.1016/j.biombioe.2020.105927.
-
Park H, Jung D, Lee J, Kim P, Cho Y, Jung I, Lee CG. Improvement of biomass and fatty acid productivity in ocean cultivation of Tetraselmis sp. using hypersaline medium. J Appl Phycol. 2018;30:2725–35. https://doi.org/10.1007/s10811-018-1388-3.
-
Zhang Y, Gu Z, Ren Y, Wang L, Zhang J, Liang C, Ye N. Integrating transcriptomics and metabolomics to characterize metabolic regulation to elevated CO2 in Chlamydomonas reinhardtii. Mar Biotech. 2021;23(2):255–75. https://doi.org/10.1007/s10126-021-10021-y.
-
Gao P, Guo L, Gao M, Zhao Y, Jin C, She Z. Regulation of carbon source metabolism in mixotrophic microalgae cultivation in response to light intensity variation. J Environ Manage. 2022;302:114095. https://doi.org/10.1016/j.jenvman.2021.114095.
-
Kim S, Moon M, Kwak M, Lee B, Chang YK. Statistical optimization of light intensity and CO2 concentration for lipid production derived from attached cultivation of green microalga Ettlia sp. Sci Rep. 2018;8:5390. https://doi.org/10.1038/s41598-018-33793-1.
-
Choudhary R, Rajput VD, Ghodake G, Ahmad F, Meena M, Rehman RU, Prasad R, Sharma RK, Singh R, Seth CS. Comprehensive journey from past to present to future about seed priming with hydrogen peroxide and hydrogen sulfide in relation to drought, temperature, UV and ozone stresses—a review. Plant Soil. 2024;500:351–73. https://doi.org/10.1007/s11104-024-06499-9.
-
Ferrer-Ledo N, Stegemüller L, Janssen M, Wijffels RH, Barbosa MJ. Growth and fatty acid distribution over lipid classes in Nannochloropsis oceanica acclimated to different temperatures. Front Plant Sci. 2023;14:1078998. https://doi.org/10.3389/fpls.2023.1078998.
-
Lv JM, Cheng LH, Xu XH, Zhang L, Chen HL. Enhanced lipid production of Chlorella vulgaris by adjustment of cultivation conditions. Bioresour Technol. 2010;101(17):6797–804. https://doi.org/10.1016/j.biortech.2010.03.120.
-
Calhoun S, Bell TAS, Dahlin LR, Kunde Y, LaButti K, Louie KB, Kuftin A, Treen D, Dilworth D, Mihaltcheva S, Daum C, Bowen BP, Northen TR, Guarnieri MT, Starkenburg SR, Grigoriev IV. A multi-omic characterization of temperature stress in a halotolerant Scenedesmus strain for algal biotechnology. Commun Bio. 2021;4(1):333. https://doi.org/10.1038/s42003-021-01859-y.
-
Leyva LA, Bashan Y, Mendoza A, de-Bashan LE. Accumulation fatty acids of in Chlorella vulgaris under heterotrophic conditions in relation to activity of acetyl-CoA carboxylase, temperature, and co-immobilization with Azospirillum brasilense. Naturwissenschaften. 2014;101(10):819–30. https://doi.org/10.1007/s00114-014-1223-x.
-
Aussant J, Guihéneuf F, Stengel DB. Impact of temperature on fatty acid composition and nutritional value in eight species of microalgae. Appl Microbiol Biotechnol. 2018;102(12):5279–97. https://doi.org/10.1007/s00253-018-9001-x.
-
Vidotti AD, Riaño-Pachón DM, Mattiello L, Giraldi LA, Winck FV, Franco TT. Analysis of autotrophic, mixotrophic and heterotrophic phenotypes in the microalgae Chlorella vulgaris using time-resolved proteomics and transcriptomics approaches. Algal Res. 2020;51:102060. https://doi.org/10.1016/j.algal.2020.102060.
-
Shi TQ, Wang LR, Zhang ZX, Sun XM, Huang H. Stresses as first-line tools for enhancing lipid and carotenoid production in microalgae. Front Bioeng Biotechnol. 2020;8:610. https://doi.org/10.3389/fbioe.2020.00610.
-
Sangela V, Kumar M, Choudhary S, Gour VS, Meena M, Vinayak V, Harish. Effect of nitrogen, phosphorus and sodium bicarbonate on lipid production and fatty acid profile in Coelastrella terrestris. Biocatal Agric Biotechnol. 2022;45:102518. https://doi.org/10.1016/j.bcab.2022.102518.
-
Breuer G, Lamers PP, Martens DE, Draaisma RB, Wijffels RH. The impact of nitrogen starvation on the dynamics of triacylglycerol accumulation in nine microalgae strains. Bioresour Technol. 2012;124:217–26. https://doi.org/10.1016/j.biortech.2012.08.003.
-
De Bhowmick G, Koduru L, Sen R. Metabolic pathway engineering towards enhancing microalgal lipid biosynthesis for biofuel application—a review. Renew Sust Energ Rev. 2015;50:1239–53. https://doi.org/10.1016/j.rser.2015.04.131.
-
Liang MH, Jiang JG. Advancing oleaginous microorganisms to produce lipid via metabolic engineering technology. Prog Lipid Res. 2013;52(4):395–408. https://doi.org/10.1016/j.plipres.2013.05.002.
-
Mandalam RK, Palsson B. Elemental balancing of biomass and medium composition enhances growth capacity in high-density Chlorella vulgaris cultures. Biotechnol Bioeng. 1998;59(5):605–11. https://doi.org/10.1002/(sici)1097-0290(19980905)59:5%3c605::aid-bit11%3e3.0.co;2-8.
-
Rodolfi L, Chini Zittelli G, Bassi N, Padovani G, Biondi N, Bonini G, Tredici MR. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol Bioeng. 2009;102(1):100–12. https://doi.org/10.1002/bit.22033.
-
Trivedi J, Agrawal D, Atray N, Ray A. Enhanced lipid production in Scenedesmus obliquus via nitrogen starvation in a two-stage cultivation process and evaluation for biodiesel production. Fuel. 2022;316:123418. https://doi.org/10.1016/j.fuel.2022.123418.
-
Swapnil P, Meena M, Rai AK. Molecular interaction of nitrate transporter proteins with recombinant glycinebetaine results in efficient nitrate uptake in the cyanobacterium Anabaena PCC 7120. PLoS ONE. 2021;16(11):e0257870. https://doi.org/10.1371/journal.pone.0257870.
-
Das S, Das S, Ghangrekar MM. Efficacious bioremediation of heavy metals and radionuclides from wastewater employing aquatic macro-and microphytes. J Basic Microbiol. 2022;62(3–4):260–78. https://doi.org/10.1002/jobm.202100372.
-
Yaashikaa PR, Kumar PS, Jeevanantham S, Saravanan R. A review on bioremediation approach for heavy metal detoxification and accumulation in plants. Environ Pollut. 2022;301:119035. https://doi.org/10.1016/j.envpol.2022.119035.
-
Tripathi S, Poluri KM. Metallothionein-and phytochelatin-assisted mechanism of heavy metal detoxification in microalgae. In: Hasanuzzaman M, editor. Approaches to the remediation of inorganic pollutants. Springer, Singapore; 2021. p. 323–44. https://doi.org/10.1007/978-981-15-6221-1_16.
-
Ren HY, Liu BF, Kong F, Zhao L, Xie GJ, Ren NQ. Enhanced lipid accumulation of green microalga Scenedesmus sp. by metal ions and EDTA addition. Bioresour Technol. 2014;169:763–77. https://doi.org/10.1016/j.biortech.2014.06.062.
-
Volgusheva A, Kukarskikh G, Krendeleva T, Rubin A, Mamedov F. Hydrogen photoproduction in green algae Chlamydomonas reinhardtii under magnesium deprivation. Rsc Adv. 2015;5(8):5633–7. https://doi.org/10.1039/C4RA12710B.
-
Guo H, Li T, Zhao Y, Yu X. Role of copper in the enhancement of astaxanthin and lipid coaccumulation in Haematococcus pluvialis exposed to abiotic stress conditions. Bioresour Technol. 2021;335:125265. https://doi.org/10.1016/j.biortech.2021.125265.
-
Aguilar-Ruiz RJ, Martínez-Macias MDR, Sánchez-Machado DI, López-Cervantes J, Dévora-Isiordia GE, Nateras-Ramírez O. Removal of copper improves the lipid content in Nannochloropsis oculata culture. Environ Sci Pollut Res Int. 2020;27(35):44195–204. https://doi.org/10.1007/s11356-020-10283-4.
-
Sun J, Cheng J, Yang Z, Li K, Zhou J, Cen K. Microstructures and functional groups of Nannochloropsis sp. cells with arsenic adsorption and lipid accumulation. Bioresour Technol. 2015;194:305–11. https://doi.org/10.1016/j.biortech.2015.07.041.
-
Zhao Y, Song X, Yu L, Han B, Li T, Yu X. Influence of cadmium stress on the lipid production and cadmium bioresorption by Monoraphidium sp. QLY-1. Energy Convers Manag. 2019;188:76–85. https://doi.org/10.1016/j.enconman.2019.03.041.
-
Yang J, Cao J, Xing G, Yuan H. Lipid production combined with biosorption and bioaccumulation of cadmium, copper, manganese and zinc by oleaginous microalgae Chlorella minutissima UTEX2341. Bioresour Technol. 2015;175:537–44. https://doi.org/10.1016/j.biortech.2014.10.124.
-
León-Vaz A, Romero LC, Gotor C, León R, Vigara J. Effect of cadmium in the microalga Chlorella sorokiniana: a proteomic study. Ecotoxicol Environ Saf. 2021;207:111301. https://doi.org/10.1016/j.ecoenv.2020.111301.
-
Battah M, El-Ayoty Y, Abomohra AEF, El-Ghany SA, Esmael A. Effect of Mn2+, Co2+ and H2O2 on biomass and lipids of the green microalga Chlorella vulgaris as a potential candidate for biodiesel production. Ann Microbiol. 2015;65(1):155–62. https://doi.org/10.1007/s13213-014-0846-7.
-
Swapnil P, Singh LA, Mandal C, Sahoo A, Batool F, Meena M, Kumari P, Zehra A. Functional characterization of microbes and their association with unwanted substance for wastewater treatment processes. J Water Proc Eng. 2023;54:103983. https://doi.org/10.1016/j.jwpe.2023.103983.
-
Kumawat G, Vyas P, Choudhary S, Meena M, Harish. Microalgal biodiesel as a sustainable and green energy alternative: a metabolomic approach. Biomass Bioenergy. 2024;186:107257. https://doi.org/10.1016/j.biombioe.2024.107257.
-
Aditi, Bhardwaj R, Yadav A, Swapnil P, Meena M. Characterization of microalgal β-carotene and astaxanthin: exploring their health-promoting properties under the effect of salinity and light intensity. Biotechnol Biofuels Bioproducts. 2025;18(1):18. https://doi.org/10.1186/s13068-025-02612-x.
-
Gour RS, Garlapati VK, Kant A. Effect of salinity stress on lipid accumulation in Scenedesmus sp. and Chlorella sp.: feasibility of stepwise culturing. Curr Microbiol. 2020;77(5):779–85. https://doi.org/10.1007/s00284-019-01860-z.
-
Mou Y, Liu N, Su K, Li X, Lu T, Yu Z, Song M. The growth and lipid accumulation of Scenedesmus quadricauda under nitrogen starvation stress during xylose mixotrophic/heterotrophic cultivation. Env Sci Pollut Res Int. 2022;30(44):98934–46. https://doi.org/10.1007/s11356-022-24579-0.
-
Bartley ML, Boeing WJ, Corcoran AA, Holguin FO, Schaub T. Effects of salinity on growth and lipid accumulation of biofuel microalga Nannochloropsis salina and invading organisms. Biomass Bioenergy. 2013;54:83–8. https://doi.org/10.1016/j.biombioe.2013.03.026.
-
Salama E, Kabra AN, Ji M, Kim JR, Min B, Jeon B. Enhancement of microalgae growth and fatty acid content under the influence of phytohormones. Bioresour Technol. 2014;172:97–103. https://doi.org/10.1016/j.biortech.2014.09.002.
-
Pandit PR, Fulekar MH, Karuna MSL. Effect of salinity stress on growth, lipid productivity, fatty acid composition, and biodiesel properties in Acutodesmus obliquus and Chlorella vulgaris. Environ Sci Pollut Res Int. 2017;24(15):13437–51. https://doi.org/10.1007/s11356-017-8875-y.
-
Chang W, Li Y, Qu Y, Liu Y, Zhang G, Zhao Y, Liu S. Mixotrophic cultivation of microalgae to enhance the biomass and lipid production with synergistic effect of red light and phytohormone IAA. Renew Energy. 2022;187:819–28. https://doi.org/10.1016/j.renene.2022.01.108.
-
Zhao N, Liu F, Dong W, Yu J, Halverson LJ, Xie B. Quantitative proteomics insights into Chlamydomonas reinhardtii thermal tolerance enhancement by a mutualistic interaction with Sinorhizobium meliloti. Microbiol Spectr. 2024;12(8):e00219-24. https://doi.org/10.1128/spectrum.00219-24.
-
Samani MSM, Mansouri H. The novel strategy for enhancing growth and lipid accumulation in Chlorella vulgaris microalgae cultured in dairy wastewater by monochromatic LEDs and melatonin. J Appl Phycol. 2022;35(2):593–601. https://doi.org/10.1007/s10811-022-02898-6.
-
Khalili Z, Jalili H, Noroozi M, Amrane A, Ashtiani FR. Linoleic-acid-enhanced astaxanthin content of Chlorella sorokiniana (Chlorophyta) under normal and light shock conditions. Phycologia. 2019;59(1):54–62. https://doi.org/10.1080/00318884.2019.1670012.
-
Chen J, Wei D, Lim P. Enhanced coproduction of astaxanthin and lipids by the green microalga Chromochloris zofingiensis: selected phytohormones as positive stimulators. Bioresour Technol. 2019;295:122242. https://doi.org/10.1016/j.biortech.2019.122242.
-
Lin Y, Dai Y, Xu W, Wu X, Li Y, Zhu H, Zhou H. The growth, lipid accumulation and fatty acid profile analysis by abscisic acid and indol-3-acetic acid induced in Chlorella sp. FACHB-8. Int J Mol Sci. 2022;23(7):4064. https://doi.org/10.3390/ijms23074064.
-
Hu Q, Song M, Huang D, Hu Z, Wu Y, Wang C. Haematococcus pluvialis accumulated lipid and astaxanthin in a moderate and sustainable way by the self-protection mechanism of salicylic acid under sodium acetate stress. Front Plant Sci. 2021;12:763742. https://doi.org/10.3389/fpls.2021.763742.
-
Sivaramakrishnan R, Incharoensakdi A. Plant hormone induced enrichment of Chlorella sp. omega-3 fatty acids. Biotechnol Biofuels. 2020;13:7. https://doi.org/10.1186/s13068-019-1647-9.
-
Jiang W, Li C, Li Y, Peng H. Metabolic engineering strategies for improved lipid production and cellular physiological responses in yeast Saccharomyces cerevisiae. J Fungi. 2022;8(5):427. https://doi.org/10.3390/jof8050427.
-
Zhang J, Hao Q, Bai L, Xu J, Yin W, Song L, Xu L, Guo X, Fan C, Chen Y, Ruan J, Hao S, Li Y, Wang R, Hu Z. Overexpression of the soybean transcription factor GmDof4 significantly enhances the lipid content of Chlorella ellipsoidea. Biotechnol Biofuels. 2014;7:128. https://doi.org/10.1186/s13068-014-0128-4.
-
Brennan L, Owende P. Biofuels from microalgae—a review of technologies for production, processing, and extractions of biofuels and co-products. Renew Sust Energ Rev. 2010;14(2):557–77. https://doi.org/10.1016/j.rser.2009.10.009.
-
Mata TM, Martins AA, Caetano NS. Microalgae for biodiesel production and other applications: a review. Renew Sust Energ Rev. 2010;14(1):217–32. https://doi.org/10.1016/j.rser.2009.07.020.
-
Kang NK, Baek K, Koh HG, Atkinson CA, Ort DR, Jin YS. Microalgal metabolic engineering strategies for the production of fuels and chemicals. Bioresour Technol. 2022;345:126529. https://doi.org/10.1016/j.biortech.2021.126529.
-
Seth K, Kumar A, Rastogi RP, Meena M, Vinayak V, Harish. Bioprospecting of fucoxanthin from diatoms—challenges and perspectives. Algal Res. 2021;60:102475. https://doi.org/10.1016/j.algal.2021.102475.
-
Ferreira GF, Pinto LR, Maciel Filho R, Fregolente LV. A review on lipid production from microalgae: association between cultivation using waste streams and fatty acid profiles. Renew Sust Energ Rev. 2019;109:448–66. https://doi.org/10.1016/j.rser.2019.04.052.
-
Blatti JL, Michaud J, Burkart MD. Engineering fatty acid biosynthesis in microalgae for sustainable biodiesel. Curr Opin Chem Biol. 2013;17(3):496–505. https://doi.org/10.1016/j.cbpa.2013.04.007.
-
La Russa M, Bogen C, Uhmeyer A, Doebbe A, Filippone E, Kruse O, Mussgnug JH. Functional analysis of three type-2 DGAT homologue genes for triacylglycerol production in the green microalga Chlamydomonas reinhardtii. J Biotechnol. 2012;162(1):13–20. https://doi.org/10.1016/j.jbiotec.2012.04.006.
-
Deng XD, Gu B, Li YJ, Hu XW, Guo JC, Fei XW. The roles of acyl-CoA: diacylglycerol acyltransferase 2 genes in the biosynthesis of triacylglycerols by the green algae Chlamydomonas reinhardtii. Mol Plant. 2012;5(4):945–7. https://doi.org/10.1093/mp/sss040.
-
Haslam RP, Hamilton ML, Economou CK, Smith R, Hassall KL, Napier JA, Sayanova O. Overexpression of an endogenous type 2 diacylglycerol acyltransferase in the marine diatom Phaeodactylum tricornutum enhances lipid production and omega-3 long-chain polyunsaturated fatty acid content. Biotechnol Biofuels. 2020;13:87. https://doi.org/10.1186/s13068-020-01726-8.
-
Bellou S, Baeshen MN, Elazzazy AM, Aggeli D, Sayegh F, Aggelis G. Microalgal lipids biochemistry and biotechnological perspectives. Biotech Adv. 2014;32(8):1476–93. https://doi.org/10.1016/j.biotechadv.2014.10.003.
-
Hamilton ML, Haslam RP, Napier JA, Sayanova O. Metabolic engineering of Phaeodactylum tricornutum for the enhanced accumulation of omega-3 long chain polyunsaturated fatty acids. Metab Eng. 2014;22:3–9. https://doi.org/10.1016/j.ymben.2013.12.003.
-
Moellering ER, Miller R, Benning C. Molecular genetics of lipid metabolism in the model green alga Chlamydomonas reinhardtii. In: Wada H, Murata N, editors. Lipids in photosynthesis. Advances in photosynthesis and respiration. vol 30. Springer, Dordrecht; 2009. p. 139–55. https://doi.org/10.1007/978-90-481-2863-1_7.
-
Muñoz CF, Südfeld C, Naduthodi MI, Weusthuis RA, Barbosa MJ, Wijffels RH, D’Adamo S. Genetic engineering of microalgae for enhanced lipid production. Biotechnol Adv. 2021;52:107836. https://doi.org/10.1016/j.biotechadv.2021.107836.
-
Swapnil P, Meena M, Singh SK, Dhuldhaj UP, Marwal A. Vital roles of carotenoids in plants and humans to deteriorate stress with its structure, biosynthesis, metabolic engineering and functional aspects. Curr Plant Biol. 2021;26:100203. https://doi.org/10.1016/j.cpb.2021.100203.
-
Maoka T. Carotenoids as natural functional pigments. J Nat Med. 2020;74(1):1–16. https://doi.org/10.1007/s11418-019-01364-x.
-
Botella-Pavía P, Rodríguez-Concepción M. Carotenoid biotechnology in plants for nutritionally improved foods. Physiol Plant. 2006;126(3):369–81. https://doi.org/10.1111/j.1399-3054.2006.00632.x.
-
Lichtenthaler HK. The 1-deoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Ann Rev Plant Biol. 1999;50(1):47–65. https://doi.org/10.1146/annurev.arplant.50.1.47.
-
Niklitschek M, Alcaíno J, Barahona S, Sepúlveda D, Lozano C, Carmona M, Marcoleta A, Martínez C, Lodato P, Baeza M, Cifuentes V. Genomic organization of the structural genes controlling the astaxanthin biosynthesis pathway of Xanthophyllomyces dendrorhous. Bio Res. 2008;41(1):93–108. https://doi.org/10.4067/S0716-97602008000100011.
-
Cunningham FX Jr, Gantt E. Genes and enzymes of carotenoid biosynthesis in plants. Annu Rev Plant Biol. 1998;49(1):557–83. https://doi.org/10.1146/annurev.arplant.49.1.557.
-
Lee P, Schmidt-Dannert C. Metabolic engineering towards biotechnological production of carotenoids in microorganisms. Appl Microbiol Biotechnol. 2002;60:1–11. https://doi.org/10.1007/s00253-002-1101-x.
-
Britton G. Structure and properties of carotenoids in relation to function. FASEB J. 1995;9(15):1551–8. https://doi.org/10.1096/fasebj.9.15.8529834.
-
Cunningham FX Jr, Pogson B, Sun Z, McDonald KA, DellaPenna D, Gantt E. Functional analysis of the beta and epsilon lycopene cyclase enzymes of Arabidopsis reveals a mechanism for control of cyclic carotenoid formation. Plant Cell. 1996;8(9):1613–26. https://doi.org/10.1105/tpc.8.9.1613.
-
Liu S, Liu B, Wang H, Xiao S, Li Y, Wang J. Production of astaxanthin at moderate temperature in Xanthophyllomyces dendrorhous using a two-step process. Eng Life Sci. 2018;18(10):706–10. https://doi.org/10.1002/elsc.201800065.
-
Tran HD, Do TT, Le TL, Nguyen MLT, Pham CH, Melkonian M. Cultivation of Haematococcus pluvialis for astaxanthin production on angled bench-scale and large-scale biofilm-based photobioreactors. Viet J Sci Technol Eng. 2019;61(3):61–70. https://doi.org/10.31276/VJSTE.61(3).61-70.
-
Le-Feuvre R, Moraga-Suazo P, Gonzalez J, Martin SS, Henríquez V, Donoso A, Agurto-Muñoz C. Biotechnology applied to Haematococcus pluvialis Fotow: challenges and prospects for the enhancement of astaxanthin accumulation. J Appl Phycol. 2020;32:3831–52. https://doi.org/10.1007/s10811-020-02231-z.
-
Zhao W, Ioannidis MA. Gas exsolution and flow during supersaturated water injection in porous media: I. Pore network modeling. Adv Water Resour. 2011;34(1):2–14. https://doi.org/10.1016/j.advwatres.2010.09.010.
-
Mansouri H, Nezhad SE, Kamyab H, Chelliapan S, Kirpichnikova I. The effects of aeration and mixotrophy by acetate and pyruvate on the growth parameters in Scenedesmus obliquus. Biomass Convers Biorefinery. 2022;12(10):4611–20. https://doi.org/10.1007/s13399-022-02676-x.
-
Magdaong JB, Ubando AT, Culaba AB, Chang JS, Chen WH. Effect of aeration rate and light cycle on the growth characteristics of Chlorella sorokiniana in a photobioreactor. IOP Confer Ser Earth Environ Sci. 2019;268(1):012112. https://doi.org/10.1088/1755-1315/268/1/012112.
-
Goswami RK, Agrawal K, Mehariya S, Rajagopal R, Karthikeyan OP, Verma P. Development of economical and sustainable cultivation system for biomass production and simultaneous treatment of municipal wastewater using Tetraselmis indica BDUG001. Environ Technol. 2023;1–21. https://doi.org/10.1080/09593330.2023.2166429.
-
Mehariya S, Goswami RK, Verma P, Lavecchia R, Zuorro A. Integrated approach for wastewater treatment and biofuel production in microalgae biorefineries. Energies. 2021;14(8):2282. https://doi.org/10.3390/en14082282.
-
Goswami RK, Mehariya S, Karthikeyan OP, Verma P. Fungi-assisted bio-flocculation of Picochlorum sp.: a novel bio-assisted treatment system for municipal wastewater. J Water Process Eng. 2023;57:104666. https://doi.org/10.1016/j.jwpe.2023.104666.
-
Oslan SNH, Oslan SN, Mohamad R, Tan JS, Yusoff AH, Matanjun P, Huda N. Bioprocess strategy of Haematococcus lacustris for biomass and astaxanthin production keys to commercialization: perspective and future direction. Fermentation. 2022;8(4):179. https://doi.org/10.3390/fermentation8040179.
-
Radice RP, Fiorentino R, De Luca M, Limongi AR, Viviano E, Bermano G, Martelli G. An innovative protocol to select the best growth phase for astaxanthin biosynthesis in H. pluvialis. Biotechnol Rep. 2021;31:e00655. https://doi.org/10.1016/j.btre.2021.e00655
-
Bhattacharya T, Rather G, Akter R, Kabir MT, Rauf A, Rahman M. Nutraceuticals and bio-inspired materials from microalgae and their future perspectives. Curr Top Med Chem. 2021;21(12):1037–51. https://doi.org/10.2174/1568026621666210524095925.
-
Kumar M, Sabu S, Sangela V, Meena M, Rajput VD, Minkina T, Vinayak V, Harish. The mechanism of nanoparticle toxicity to cyanobacteria. Arch Microbiol. 2023;205:30. https://doi.org/10.1007/s00203-022-03370-2
-
Kumar S, Masurkar P, Sravani B, Bag D, Sharma KR, Singh P, Korra T, Meena M, Swapnil P, Rajput VD, Minkina T. A review on phytotoxicity and defense mechanism of silver nanoparticles (AgNPs) on plants. J Nanopart Res. 2023;25:54. https://doi.org/10.1007/s11051-023-05708-3.
-
Mariyam S, Upadhyay SK, Chakraborty K, Verma KK, Duhan JS, Muneer S, Meena M, Sharma RK, Ghodake G, Seth CS. Nanotechnology, a frontier in agricultural science, a novel approach in abiotic stress management and convergence with new age medicine—a review. Sci Total Environ. 2023;912:169097. https://doi.org/10.1016/j.scitotenv.2023.169097.
-
Nasri N, Keyhanfar M, Behbahani M, Dini G. Enhancement of astaxanthin production in Haematococcus pluvialis using zinc oxide nanoparticles. J Biotechnol. 2021;342:72–8. https://doi.org/10.1016/j.jbiotec.2021.10.004.
-
Patel AK, Tambat VS, Chen CW, Chauhan AS, Kumar P, Vadrale AP, Singhania RR. Recent advancements in astaxanthin production from microalgae: a review. Bioresour Technol. 2022;364:128030. https://doi.org/10.1016/j.biortech.2022.128030.
-
Venckus P, Endriukaitytė I, Čekuolytė K, Gudiukaitė R, Pakalniškis A, Lastauskienė E. Effect of biosynthesized silver nanoparticles on the growth of the green microalga Haematococcus pluvialis and astaxanthin synthesis. Nanomaterials. 2023;13(10):1618. https://doi.org/10.3390/nano13101618.
-
Wang X, Song Y, Liu B, Hang W, Li R, Cui H, Li R, Jia X. Enhancement of astaxanthin biosynthesis in Haematococcus pluvialis via inhibition of autophagy by 3-methyladenine under high light. Algal Res. 2020;50:101991. https://doi.org/10.1016/j.algal.2020.101991.
-
Xiong X, Liu W, Jiang J, Xu L, Huang L, Cao J. Efficient genome editing of Brassica campestris based on the CRISPR/Cas9 system. Mol Genet Genomics. 2019;294(5):1251–61. https://doi.org/10.1007/s00438-019-01564-w.
-
Han D, Li Y, Hu Q. Astaxanthin in microalgae: pathways, functions, and biotechnological implications. Algae. 2013;28(2):131–47. https://doi.org/10.4490/algae.2013.28.2.131.
-
Indrayani I, Egeland ES, Moheimani NR, Borowitzka MA. Carotenoid production of Botryococcus braunii CCAP 807/2 under different growth conditions. J Appl Phycol. 2022;34(3):1177–88. https://doi.org/10.1007/s10811-022-02682-6.
-
Vieira ARL. Identification of microalgae strains producing high-value compounds. Instituto de Ciências Biomédicas Abel Salazar da Universidade do Porto, 2022.
-
Barati Aburai N, Sumida D, Abe K. Effect of light level and salinity on the composition and accumulation of free and ester-type carotenoids in the aerial microalga Scenedesmus sp. (Chlorophyceae). Algal Res. 2015;8:30–6. https://doi.org/10.1016/j.algal.2015.01.005.
-
Peng J, Yuan JP, Wang JH. Effect of diets supplemented with different sources of astaxanthin on the gonad of the sea urchin Anthocidaris crassispina. Nutrients. 2012;4(8):922–34. https://doi.org/10.3390/nu4080922.
-
Park EK, Lee CG. Astaxanthin production by Haematococcus pluvialis under various light intensities and wavelengths. J Microbiol Biotechnol. 2001;11(6):1024–30.
-
Orosa M, Valero JF, Herrero C, Abalde J. Comparison of the accumulation of astaxanthin in Haematococcus pluvialis and other green microalgae under N-starvation and high light conditions. Biotechnol Lett. 2001;23:1079–85. https://doi.org/10.1023/A:1010510508384.
-
Lv R, Liu K, Chen F, Xing H, Xu N, Sun X, Hu C. Buffering culture solution significantly improves astaxanthin production efficiency of mixotrophic Haematococcus pluvialis. Bioresour Technol. 2022;354:127175. https://doi.org/10.1016/j.biortech.2022.127175.
-
Yuan J, Chen F. Purification of trans-astaxanthin from a high-yielding astaxanthin ester-producing strain of the microalga Haematococcus pluvialis. Food Chem. 2000;68(4):443–8. https://doi.org/10.1016/s0308-8146(99)00219-8.
-
Holtin K, Kuehnle M, Rehbein J, Schuler P, Nicholson G, Albert K. Determination of astaxanthin and astaxanthin esters in the microalgae Haematococcus pluvialis by LC-(APCI)MS and characterization of predominant carotenoid isomers by NMR spectroscopy. Anal Bioanal Chem. 2009;395(6):1613–22. https://doi.org/10.1007/s00216-009-2837-2.
-
Chen G, Wang B, Han D, Sommerfeld M, Lu Y, Chen F, Hu Q. Molecular mechanisms of the coordination between astaxanthin and fatty acid biosynthesis in Haematococcus pluvialis (Chlorophyceae). Plant J. 2014;81(1):95–107. https://doi.org/10.1111/tpj.12713.
-
Grünewald K, Hirschberg J, Hagen C. Ketocarotenoid biosynthesis outside of plastids in the unicellular green alga Haematococcus pluvialis. J Biol Chem. 2001;276(8):6023–9. https://doi.org/10.1074/jbc.m006400200.
-
Collins AM, Jones HDT, Han D, Hu Q, Beechem TE, Timlin JA. Carotenoid distribution in living cells of Haematococcus pluvialis (Chlorophyceae). PLoS ONE. 2011;6(9):e24302. https://doi.org/10.1371/journal.pone.0024302.
-
Gao Z, Li Y, Wu G, Li G, Sun H, Deng S, Shen Y, Chen G, Zhang R, Meng C, Zhang X. Transcriptome analysis in Haematococcus pluvialis: astaxanthin induction by salicylic acid (SA) and jasmonic acid (JA). PLoS ONE. 2015;10(10):e0140609. https://doi.org/10.1371/journal.pone.0140609.
-
Yen CE, Monetti M, Burri BJ, Farese RV. The triacylglycerol synthesis enzyme DGAT1 also catalyzes the synthesis of diacylglycerols, waxes, and retinyl esters. J Lipid Res. 2005;46(7):1502–11. https://doi.org/10.1194/jlr.m500036-jlr200.
-
Biester E, Hellenbrand J, Frentzen M. Multifunctional acyltransferases from Tetrahymena thermophila. Lipids. 2011;47(4):371–81. https://doi.org/10.1007/s11745-011-3642-1.
-
Chen JE, Smith AG. A look at diacylglycerol acyltransferases (DGATs) in algae. J Biotechnol. 2012;162(1):28–39. https://doi.org/10.1016/j.jbiotec.2012.05.009.
-
Li J, Han D, Wang D, Ning K, Jia J, Wei L, Jing X, Huang S, Chen J, Li Y, Hu Q, Xu J. Choreography of transcriptomes and lipidomes of Nannochloropsis reveals the mechanisms of oil synthesis in microalgae. Plant Cell. 2014;26(4):1645–65. https://doi.org/10.1105/tpc.113.121418.
-
Kwan TA, Kwan SE, Peccia J, Zimmerman JB. Selectively biorefining astaxanthin and triacylglycerol co-products from microalgae with supercritical carbon dioxide extraction. Bioresour Technol. 2018;269:81–8. https://doi.org/10.1016/j.biortech.2018.08.081.
-
Thomassen G, Van Dael M, Van Passel S. The potential of microalgae biorefineries in Belgium and India: an environmental techno-economic assessment. Bioresour Technol. 2018;267:271–80. https://doi.org/10.1016/j.biortech.2018.07.037.
-
Salama E, Kurade MB, Abou-Shanab RA, El-Dalatony MM, Yang I, Min B, Jeon B. Recent progress in microalgal biomass production coupled with wastewater treatment for biofuel generation. Renew Sust Energ Rev. 2017;79:1189–211. https://doi.org/10.1016/j.rser.2017.05.091.
-
Chandra R, Iqbal HM, Vishal G, Lee H, Nagra S. Algal biorefinery: a sustainable approach to valorize algal-based biomass towards multiple product recovery. Bioresour Technol. 2019;278:346–59. https://doi.org/10.1016/j.biortech.2019.01.104.
-
Levasseur W, Perré P, Pozzobon V. A review of high value-added molecules production by microalgae in light of the classification. Biotechnol Adv. 2020;41:107545. https://doi.org/10.1016/j.biotechadv.2020.107545.
-
Liyanaarachchi VC, Premaratne M, Ariyadasa TU, Nimarshana P, Malik A. Two-stage cultivation of microalgae for production of high-value compounds and biofuels: a review. Algal Res. 2021;57:102353. https://doi.org/10.1016/j.algal.2021.102353.
-
Yu C, Li X, Han B, Zhao Y, Geng S, Ning D, Ma T, Yu X. Simultaneous improvement of astaxanthin and lipid production of Haematococcus pluvialis by using walnut shell extracts. Algal Res. 2020;54:102171. https://doi.org/10.1016/j.algal.2020.102171.
-
Nishshanka GKSH, Liyanaarachchi VC, Premaratne M, Nimarshana PHV, Ariyadasa TU, Kornaros M. Wastewater-based microalgal biorefineries for the production of astaxanthin and co-products: current status, challenges and future perspectives. Bioresour Technol. 2021;342:126018. https://doi.org/10.1016/j.biortech.2021.126018.
-
Siddiki SYA, Mofijur M, Kumar PS, Ahmed SF, Inayat A, Kusumo F, Badruddin IA, Yunus Khan TM, Nghiem LD, Ong HC, Mahlia TMI. Microalgae biomass as a sustainable source for biofuel, biochemical and biobased value-added products: an integrated biorefinery concept. Fuel. 2022;307:121782. https://doi.org/10.1016/j.fuel.2021.121782.
-
Rafa N, Ahmed SF, Badruddin IA, Mofijur M, Kamangar S. Strategies to produce cost-effective third-generation biofuel from microalgae. Front Energy Res. 2021;9:749968. https://doi.org/10.3389/fenrg.2021.749968.
-
Solovchenko AE. Recent breakthroughs in the biology of astaxanthin accumulation by microalgal cell. Photosyn Res. 2015;125(3):437–49. https://doi.org/10.1007/s11120-015-0156-3.
-
Li Q, Zhao Y, Ding W, Han B, Geng S, Ning D, Ma T, Yu X. Gamma-aminobutyric acid facilitates the simultaneous production of biomass, astaxanthin and lipids in Haematococcus pluvialis under salinity and high-light stress conditions. Bioresour Technol. 2020;320:124418. https://doi.org/10.1016/j.biortech.2020.124418.
-
Bart JCJ, Palmeri N, Cavallaro S. Emerging new energy crops for biodiesel production. In: Bart JCJ, Palmeri N, Cavallaro S, editors. Biodiesel science and technology. Woodhead Publishing; 2010, p. 226–84. https://doi.org/10.1533/9781845697761.226
-
Sun X, Ren L, Zhao Q, Ji X, Huang H. Microalgae for the production of lipid and carotenoids: a review with focus on stress regulation and adaptation. Biotechnol Biofuels. 2018;11(1):272. https://doi.org/10.1186/s13068-018-1275-9.
-
Wang T, Zhu L, Mei L, Kanda H. Extraction and separation of natural products from microalgae and other natural sources using liquefied dimethyl ether, a green solvent: a review. Foods. 2024;13(2):352. https://doi.org/10.3390/foods13020352.
-
Cheng J, Li K, Zhu Y, Yang W, Zhou J, Cen K. Transcriptome sequencing and metabolic pathways of astaxanthin accumulated in Haematococcus pluvialis mutant under 15% CO2. Bioresour Technol. 2016;228:99–105. https://doi.org/10.1016/j.biortech.2016.12.084.
-
Zhang L, Zhang C, Liu J, Yang N. A strategy for stimulating astaxanthin and lipid production in Haematococcus pluvialis by exogenous glycerol application under low light. Algal Res. 2019;46:101779. https://doi.org/10.1016/j.algal.2019.101779.
-
Bortesi L, Fischer R. The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol Adv. 2015;33(1):41–52. https://doi.org/10.1016/j.biotechadv.2014.12.006.
-
Jeon S, Lim JM, Lee HG, Shin SE, Kang NK, Park YI, Chang YK. Current status and perspectives of genome editing technology for microalgae. Biotechnol Biofuels. 2017;10(1):267. https://doi.org/10.1186/s13068-017-0957-z.
-
Raper A, Stephenson A, Suo Z. Sharpening the scissors: mechanistic details of CRISPR/CAS9 improve functional understanding and inspire future research. J Am Chem Soc. 2018;140(36):11142–52. https://doi.org/10.1021/jas.8b05469.
-
Belhaj K, Chaparro-Garcia A, Kamoun S, Patron N, Nekrasov V. Editing plant genomes with CRISPR/Cas9. Curr Opin Biotechnol. 2015;32:76–84. https://doi.org/10.1016/j.copbio.2014.11.007.
-
Gaj T, Gersbach CA, Barbas CF. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013;31(7):397–405. https://doi.org/10.1016/j.tibtech.2013.04.004.
-
Mahfouz MM, Piatek A, Stewart CN Jr. Genome engineering via TALENs and CRISPR/Cas9 systems: challenges and perspectives. Plant Biotechnol J. 2014;12(8):1006–14. https://doi.org/10.1111/pbi.12256.
-
Zhang F, Maeder ML, Unger-Wallace E, Hoshaw JP, Reyon D, Christian M, Voytas DF. High frequency targeted mutagenesis in Arabidopsis thaliana using zinc finger nucleases. Proc Natl Acad Sci USA. 2010;107(26):12028–33. https://doi.org/10.1073/pnas.0914991107.
-
Wang X, Xiong X, Sa N, Roje S, Chen S. Metabolic engineering of enhanced glycerol-3-phosphate synthesis to increase lipid production in Synechocystis sp. PCC 6803. Appl Microbiol Biotechnol. 2016;100:6091–101. https://doi.org/10.1007/s00253-016-7521-9.
-
Hsu PD, Lander ES, Zhang F. Development and applications of CRISPR-Cas9 for genome engineering. Cell. 2014;157(6):1262–78. https://doi.org/10.1016/j.cell.2014.05.010.
-
Zhang Z, Mao Y, Ha S, Liu W, Botella JR, Zhu JK. A multiplex CRISPR/Cas9 platform for fast and efficient editing of multiple genes in Arabidopsis. Plant Cell Rep. 2016;35(7):1519–33. https://doi.org/10.1007/s00299-015-1900-z.
-
Serif M, Dubois G, Finoux AL, Teste MA, Jallet D, Daboussi F. One-step generation of multiple gene knock-outs in the diatom Phaeodactylum tricornutum by DNA-free genome editing. Nat Commun. 2018;9(1):3924. https://doi.org/10.1038/s41467-018-06378-9.
-
Wang SW, Gao C, Zheng YM, Yi L, Lu JC, Huang XY, Ke AW. Current applications and future perspective of CRISPR/Cas9 gene editing in cancer. Mol Cancer. 2022;21(1):57. https://doi.org/10.1186/s12943-022-01518-8.
-
Asmamaw M, Zawdie B. Mechanism and applications of CRISPR/Cas-9-mediated genome editing. Biol Targets Ther. 2021;15:353–61. https://doi.org/10.2147/BTT.S326422.
-
Nelson DR, Hazzouri KM, Lauersen KJ, Jaiswal A, Chaiboonchoe A, Mystikou A, Fu W, Daakour S, Dohai B, Alzahmi A, Nobles D, Hurd M, Sexton J, Preston MJ, Blanchette J, Lomas MW, Amiri KM, Salehi-Ashtiani K. Large-scale genome sequencing reveals the driving forces of viruses in microalgal evolution. Cell Host Microbe. 2021;29(2):250–66. https://doi.org/10.1016/j.chom.2020.12.005.
-
Clement K, Hsu JY, Canver MC, Joung JK, Pinello L. Technologies and computational analysis strategies for CRISPR applications. Mol Cell. 2020;79(1):11–29. https://doi.org/10.1016/j.molcel.2020.06.012.
-
Modrzejewski D, Hartung F, Lehnert H, Sprink T, Kohl C, Keilwagen J, Wilhelm R. Which factors affect the occurrence of off-target effects caused by the use of CRISPR/Cas: a systematic review in plants. Front Plant Sci. 2020;11:574959. https://doi.org/10.3389/fpls.2020.574959.
-
Přibylová A, Fischer L, Pyott DE, Bassett A, Molnar A. DNA methylation can alter CRISPR/Cas9 editing frequency and DNA repair outcome in a target-specific manner. New Phytol. 2022;235(6):2285–99. https://doi.org/10.1111/nph.18212.
-
Sledzinski P, Nowaczyk M, Olejniczak M. Computational tools and resources supporting CRISPR-Cas experiments. Cells. 2020;9(5):1288. https://doi.org/10.3390/cells9051288.
-
Bae S, Park J, Kim J. Cas-OFFinder: a fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics. 2014;30(10):1473–5. https://doi.org/10.1093/bioinformatics/btu048.
-
Park J, Lim K, Kim J, Bae S. Cas-analyzer: an online tool for assessing genome editing results using NGS data. Bioinformatics. 2016;33(2):286–8. https://doi.org/10.1093/bioinformatics/btw561.
-
Quan J, Langelier C, Kuchta A, Batson J, Teyssier N, Lyden A, Caldera S, McGeever A, Dimitrov B, King R, Wilheim J, Murphy M, Ares LP, Travisano KA, Sit R, Amato R, Mumbengegwi DR, Smith JL, Bennett A, Gosling R, Mourani PM, Calfee CS, Neff NF, Chow ED, Kim PS, Greenhouse B, DeRisi JL, Crawford ED. FLASH: a next-generation CRISPR diagnostic for multiplexed detection of antimicrobial resistance sequences. Nucleic Acids Res. 2019;47(14):e83. https://doi.org/10.1093/nar/gkz418.
-
Lin J, Wong K. Off-target predictions in CRISPR-Cas9 gene editing using deep learning. Bioinformatics. 2018;34(17):i656–63. https://doi.org/10.1093/bioinformatics/bty554.
-
Chuai G, Ma H, Yan J, Chen M, Hong N, Xue D, Zhou C, Zhu C, Chen K, Duan B, Gu F, Qu S, Huang D, Wei J, Liu Q. DeepCRISPR: optimized CRISPR guide RNA design by deep learning. Genome Biol. 2018;19(1):80. https://doi.org/10.1186/s13059-018-1459-4.
-
Zhang W, Wang J, Wang J, Liu T. Attached cultivation of Haematococcus pluvialis for astaxanthin production. Bioresour Technol. 2014;158:329–35. https://doi.org/10.1016/j.biortech.2014.02.044.
-
Daboussi F, Leduc S, Maréchal A, Dubois G, Guyot V, Perez-Michaut C, Amato A, Falciatore A, Juillerat A, Beurdeley M, Voytas DF, Cavarec L, Duchateau P. Genome engineering empowers the diatom Phaeodactylum tricornutum for biotechnology. Nat Commun. 2014;5:3831. https://doi.org/10.1038/ncomms4831.
-
Allemailem KS, Almatroodi SA, Almatroudi A, Alrumaihi F, Abdulmonem WA, Al-Megrin WAI, Aljamaan AN, Rahmani AH, Khan AA. Recent advances in genome-editing technology with CRISPR/Cas9 variants and stimuli-responsive targeting approaches within tumor cells: a future perspective of cancer management. Int J Mol Sci. 2023;24(8):7052. https://doi.org/10.3390/ijms24087052.
-
Idoko-Akoh A, Taylor L, Sang HM, McGrew MJ. High fidelity CRISPR/Cas9 increases precise monoallelic and biallelic editing events in primordial germ cells. Sci Rep. 2018;8(1):15126. https://doi.org/10.1038/s41598-018-33244-x.
-
Torella L, Klermund J, Bilbao-Arribas M, Tamayo I, Andrieux G, Chmielewski KO, Vales A, Olagüe C, Moreno-Luqui D, Raimondi I, Abad A, Torrens-Baile J, Salido E, Huarte M, Hernaez M, Boerries M, Cathomen T, Zabaleta N, Gonzalez-Aseguinolaza G. Efficient and safe therapeutic use of paired Cas9-nickases for primary hyperoxaluria type 1. EMBO Mol Med. 2024;16(1):112–31. https://doi.org/10.1038/s44321-023-00008-8.
-
Matsumoto D, Matsugi E, Kishi K, Inoue Y, Nigorikawa K, Nomura W. SpCas9-HF1 enhances accuracy of cell cycle-dependent genome editing by increasing HDR efficiency, and by reducing off-target effects and indel rates. Mol Ther Nucleic Acids. 2024;35(1):102124. https://doi.org/10.1016/j.omtn.2024.102124.
-
Kantor A, McClements M, MacLaren R. CRISPR-Cas9 DNA base-editing and prime-editing. Int J Mol Sci. 2020;21(17):6240. https://doi.org/10.3390/ijms21176240.
-
Saha S, Maji S, Ghosh SK, Maiti MK. Engineered Chlorella vulgaris improves bioethanol production and promises prebiotic application. World J Microbiol Biotechnol. 2024;40(9):271. https://doi.org/10.1007/s11274-024-04074-z.
-
Singh RP, Yadav P, Kumar I, Solanki MK, Roychowdhury R, Kumar A, Gupta RK. Advancement of abiotic stresses for microalgal lipid production and its bioprospecting into sustainable biofuels. Sustainability. 2023;15(18):13678. https://doi.org/10.3390/su151813678.
-
Acheampong A, Li L, Elsherbiny SM, Wu Y, Swallah MS, Bondzie-Quaye P, Huang Q. A crosswalk on the genetic and conventional strategies for enhancing astaxanthin production in Haematococcus pluvialis. Crit Rev Biotechnol. 2023;44(6):1018–39. https://doi.org/10.1080/07388551.2023.2240009.
Acknowledgements
The author P.S. gratefully acknowledges the financial support provided by the funding agency Science and Engineering Research Board (SERB), Empowerment and Equity Opportunities for Excellence in Science (File Number: EEQ/2022/000455). The author P.S. gratefully acknowledges the financial support provided by the funding agency DST-FIST (GP193). The author M.M. also gratefully acknowledges the financial support provided by the funding agency Science and Engineering Research Board (SERB), State University Research Excellence (SURE) (File Number: SUR/2022/005216) for this study. The author M.M. is highly thankful to the Ministry of Education and SPD-RUSA Rajasthan for the financial support received under the RUSA-2.0 project. All the authors read and approved the content of the manuscript for publication. The authors are also grateful to their respective universities for providing support during the work. All the authors read and approve the content of the manuscript for publication.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Ethics declarations
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yadav, A., Sharma, S., Nitesh et al. Tapping the microalgal potential: genetic precision and stress-induction for enhanced astaxanthin and biofuel production. Biotechnol. Biofuels Bioprod. 18, 92 (2025). https://doi.org/10.1186/s13068-025-02656-z
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1186/s13068-025-02656-z