
Introduction
Brucellosis is a severe systemic zoonotic infection caused by various Gram-negative Brucellaspecies, including Brucella melitensis, Brucella abortus, Brucellasuis, and Brucellacanis. It predominantly affects domestic and wild animals, resulting in significant economic losses in livestock and posing a risk of human infection through various exposure routes1,2. B. melitensisis the primary and most severe causative agent of human brucellosis, capable of progressing to a chronic granulomatous stage and leading to complications such as endocarditis, osteoarthritis, meningitis, spondylitis, arthritis, and neurological disorders3,4. According to the World Health Organization (WHO), more than 500,000 new cases of brucellosis are reported worldwide annually, presenting significant challenges in the Mediterranean region, Western Asia, and select areas of Africa and Latin America5. In recent years, brucellosis cases have exhibited characteristics different from those observed a decade ago, with emerging trends including the spread from rural to urban areas, rapid geographic expansion, travel-related transmission, and persistent presence in various wild animal populations, thereby presenting increased challenges in disease management6. The diagnosis of human brucellosis remains challenging due to its non-specific clinical features, the slow growth rate in cultures, and the complexity of serodiagnosis, leading to an estimated four undetected cases for every diagnosed case7. Due to considerable economic losses in livestock, potential zoonotic transmission, and limitations in diagnosis and epidemiology, efficacious B. melitensis vaccines are urgently needed for the control and management of the disease.
The live attenuated vaccines have been the cornerstone of brucellosis control in animals, with extensive immunization programs that have dramatically reduced brucellosis prevalence, allowing some countries to even declare eradication of the disease8,9. However, despite widespread adoption, a resurgence of brucellosis has been observed recently, particularly in developing nations10. This resurgence is likely attributable to limitations inherent to these vaccines, such as residual virulence posing a risk of reversion to more virulent strains, potential side effects in pregnant animals, and interference with serological diagnostic tests10. This underscores the urgent need to develop novel, safer, and more efficacious brucellosis vaccine candidates, particularly for human application, to support current disease control programs. Despite promising preclinical results, no authorized subunit vaccine against B. melitensis infection has been approved8,9. This underscores the imperative of identifying optimal immunogenicsubunit candidates and utilizing adjuvants or delivery systems to enhance their stability and immunogenicity, thereby facilitating the effective induction of protective humoral and cellular immune responses11,12.
BMEI1873, an ortholog of B. melitensis BtaE, belongs to the Brucella trimeric autotransporter (BTAs) family. This family comprises an N-terminal signal peptide, a globular head, a connector, a stalk passenger domain, and a YadA-like anchor domain at the C-terminal13,14,15. BtaE contributes to the adhesive properties of the Brucella cell surface, facilitating bacterial adhesion to host cells and extracellular matrix (ECM) components. This mechanism aids in evading rapid clearance from host cells and enhances the pathogen’s ability to establish infection in various tissues such as lymph nodes, spleen, liver, bone marrow, mammary glands, and sex organs by macrophages16. BtaE is essential for full virulence in B. suis and mediates the adhesion of the bacterium to both hyaluronic acid and epithelial cells, with BtaE mutants displaying reduced adhesion to human cervical adenocarcinoma epithelial cells (HeLa) and human lung carcinoma epithelial cells (A549) cells17. BtaF, another member of the BTAs family in B. suis, contributes to resistance against porcine serum and bacterial adhesion to epithelial cells18,19. Furthermore, it is crucial for establishing complete virulence during the early stages of infection in a murine model. Moreover, nasal immunization with BtaF, when combined with the c-di-AMP adjuvant, protects against intragastric challenge with B. suis through the induction of potent specific antibodies and the generation of central memory CD4+ T cells18. BTAsplay a crucial role in mediating the adhesion and virulence of Brucella species, thereby highlighting the significance of clarifying their functions in the context of developing efficacious vaccines and therapeutics for brucellosis. These observations underscore the importance of augmenting the effectiveness of BTAs through their combination with adjuvants or protein carriers, which can further modulate the immune response towards TH1 polarization against Brucella species.
Lpp-OmpA is a well-established and efficient system for displaying heterologous proteins on the surface of Escherichia coli. It comprises a signal peptide and the initial nine N-terminal amino acids of Braun’s lipoprotein (Lpp), which, when fused with five of the eight membrane-spanning segments of the outer membrane protein A (OmpA), ensures the accurate insertion and anchoring of the displayed protein within the outer membrane of E. coli19,20,21. Previous studies have demonstrated that the Lpp-OmpA platform enables the presentation of various polypeptides, including beta-lactamase, cellulase, and single-chain variable fragment (scFv) antibodies on the outer membrane22. The Lpp-OmpA system has also demonstrated potential for developing live bacterial vaccines by displaying outer membrane proteins, like protein E from Haemophilus influenzae, on the surface of E. coli23. The Lpp-OmpA chimera platform was employed to present the SARS-CoV-2 S1 protein on the membrane of synthetic bacterial vesicles (SyBV), leading to the elicitation of S1-specific antibodies and T-cell responses24. Recent research has further revealed that intranasal immunization of mice with outer membrane vesicles (OMVs) decorated with the Lpp-OmpA-SARS-CoV-2 receptor-binding domain elicits a robust neutralizing antibody response against the spike protein25.
This study aimed to evaluate the potential of incorporating BtaE into the Lpp-OmpA system to augment the Th1-type immune response and confer protection against B. melitensis infection in a murine model.
Results
In Silico design and characterization of Lpp-OmpA-BtaE fusion protein
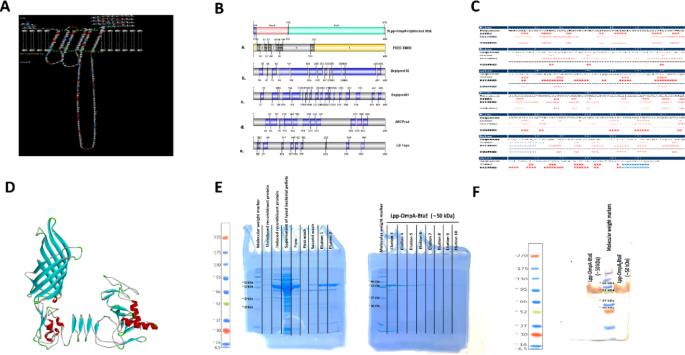
Identifying B-cell epitopes for vaccine development often relies on the pathogen’s surface-exposed proteins. Therefore, computational modeling was employed to predict the structural features of the Lpp-OmpA-BtaE fusion protein, particularly focusing on solvent-accessible loops suitable for epitope display. Bepipred, LB Tope, and other bioinformatic tools were integrated with the predicted topology of the Lpp-OmpA-BtaE fusion protein derived from PRED-TMBB analysis. This combined approach facilitated the identification of potential B-cell epitopes located within external loops at positions 27–41, 67–83, and 109–209 (Fig. 1A–C, Table 1). For major histocompatibility complex class I (MHC-I) binding, individual allele predictions identified two peptides, “WTFDKEDYSPIGDY” at positions 137–146 and “SKDSTDAVNGSQLY” at positions 341–355 of BtaE within the Lpp-OmpA-BtaE fusion protein, with scores of 0.202054 and 0.170686, respectively, suggesting potential binding to human leukocyte antigen (HLA)-A01:01. Regarding MHC-II binding, the peptide “IGDYFGNTAPADKDS” at positions 147–161 exhibited significant binding affinity to HLA-DRB302:02 (Score = 0.9551), while “GNTFLDTNGLVITGG” at positions 419–433 showed strong affinity for HLA-DRB107:01 (Score = 0.9494). Furthermore, “SQLYAVAEQATLVDG” at positions 352–366 displayed notable binding affinity to HLA-DRB1*09:01 (Score = 0.8957). Secondary structure analysis using SYMPRED, PSIPRED, and SSPro confirmed the linear and discontinuous nature of these loops, further validated by ElliPro software (Fig. 1B). Sequence homology analysis revealed a high degree of similarity (99%) between a portion of the predicted structural sequence and a B. melitensis cell surface protein (Accession No: AAL53054.1), suggesting potential antigenic cross-reactivity. Robetta’s output was chosen for the initial structural model, followed by refinement using 3DRefine to improve the QMEAN score without significantly altering the Ramachandran plot. The final three-dimensional structure displayed good agreement with the predicted secondary structure and topology (Fig. 1D). Following in silico analysis, SDS-PAGE confirmed the expression and purification of the Lpp-OmpA-BtaE fusion protein at the expected molecular weight (~ 50 kDa) (Fig. 1E). Western blot analysis using a monoclonal anti-His tag antibody further verified the identity of the purified protein (Fig. 1F).

Design and Characterization of the Lpp-OmpA-BtaE fusion protein. (A) Predicted transmembrane topology of the Lpp-OmpA-BtaEfusion protein.(B) Schematic representation of the topology (a) as predicted by PRED-TMBB2 and the linear B-cell epitopes identified by (b) Bepipred 2.0, (c) Bepipred 1.0, (d) ABCPred, and (e) LB Tope tools. (C) Predicted secondary structure of the Lpp-OmpA-BtaE fusion protein. H: helix; E: extended strand; C: coil. (D) The three-dimensional structure of the Lpp-OmpA-BtaE fusion protein was modeled by Robettaand refined using Galaxy Refine. (E) SDS-PAGE analysis of purified recombinant Lpp-OmpA-BtaE protein. (F) Western blot analysis showing purified recombinant Lpp-OmpA-BtaE proteins detected using an anti-His tag antibody.
Lpp-OmpA-BtaE immunization elicited antibody responses
To evaluate the humoral responses induced by Lpp-OmpA-BtaE + FA, we conducted a comparative analysis of serum IgG levels between immunized and non-immunized mice (Fig. 2A). Lpp-OmpA-BtaE + Freund’s adjuvant (FA) immunized mice demonstrated significantly elevated IgG levels on days 14, 28, and 42 post-initial immunization compared to the control groups (P < 0.0001; Fig. 2A). Similarly, sera from Lpp-OmpA-BtaEimmunized mice exhibited significantly higher IgG levels than those from PBS-injected mice (P < 0.0001; Fig. 2A). Moreover, IgG levels in sera from Lpp-OmpA-BtaE + FA immunized mice showed a substantial increase by 1–2 fold (P < 0.0001) at four and six weeks following booster doses compared to Lpp-OmpA-BtaE immunized mice (Fig. 2A).

Lpp-OmpA-BtaE immunization elicits antibody responses. Serum IgG (A), IgG1 (B), and IgG2a (C) titers against Lpp-OmpA-BtaE were measured in immunized and non-immunized mice. Total IgG levels were assessed at days 14, 28, and 42 post-initial immunization, whereas IgG1 and IgG2a titers were measured at day 42.The data represent the mean ± SD of 5 mice per group. ****P < 0.0001.
Furthermore, we examined IgG1 and IgG2a titers to ascertain whether the Lpp-OmpA-BtaE vaccine facilitated IgG subtype class switching. The levels of both IgG1 and IgG2a in the sera of all immunized mice were significantly higher than those in the PBS-injected group (P < 0.0001; Fig. 2B,C). Notably, IgG1 and IgG2a levels in sera from Lpp-OmpA-BtaE + FA immunized mice were significantly elevated compared to other groups (P < 0.0001; Fig. 2B,C). In Lpp-OmpA-BtaE + FA immunized mice, IgG2a levels surpassed IgG1 levels (P < 0.0001). Although no significant difference was observed in IgG1 and IgG2a responses in sera from mice vaccinated with Lpp-OmpA-BtaE alone (P > 0.05), they were significantly elevated compared to those in sera from PBS-injected mice (Fig. 2B,C).
Lpp-OmpA-BtaE stimulates splenocyte cytokine production
To assess the cytokine response of splenocytes from immunized and non-immunized mice, we measured the production of IFN-γ, IL-12, IL-4, and IL-10. As shown in Fig. 3A–D, IFN-γ, IL-12, IL-4, and IL-10 concentrations in the culture supernatant of Lpp-OmpA-BtaE + FA immune mouse splenocytes were significantly higher than those in the other groups (P < 0.004). IFN-γ, IL-12, IL-4, and IL-10 levels in Lpp-OmpA-BtaE splenocytes were significantly higher than those in PBS splenocyte cultures (P < 0.01).

Cytokine production by Lpp-OmpA-BtaE-stimulated splenocytes. Spleens from immunized and non-immunized mice were collected two weeks after the final immunization and stimulated with Lpp-OmpA-BtaE. The levels of IFN-γ (A), IL-12 (B), IL-4 (C), and IL-10 (D) in the culture supernatants were measured after 72 h. The data represent the mean ± SD of 5 mice per group. *P < 0.01, **P < 0.004***P < 0.002 and ****P < 0.0001.
Lpp-OmpA-BtaE immunization enhances T cell populations
To evaluate the cellular immune response induced by Lpp-OmpA-BtaE immunization, we employed flow cytometry with specific monoclonal antibodies to quantify CD8+ and CD4+ T cell populations in splenocytes. Mice immunized with Lpp-OmpA-BtaE + FA exhibited significantly increased percentages of both CD8+ and CD4+ T cells compared to Lpp-OmpA-BtaE alone (P < 0.01; Fig. 4A-E). Furthermore, the Lpp-OmpA-BtaE group displayed a greater expansion of these T cell populations compared to the PBS control group (P < 0.05; Fig. 4A–E).

Lpp-OmpA-BtaE immunization enhances T cell populations. Flow cytometric analysis of CD8+ and CD4+ T-cells in splenocytes from mice immunized with Lpp-OmpA-BtaE + FA (A), Lpp-OmpA-BtaE (B) and PBS (C). The percentage of CD8+ (D) and CD4+ (E) T-cells collected from immunized and nonimmunized mice. The data represent the mean ± SD of 5 mice per group. *P < 0.01, **P < 0.004***P < 0.002 and ****P < 0.0001.
Lpp-OmpA-BtaE immunization reduced bacterial burden
The protective efficacy of Lpp-OmpA-BtaE immunization against B. melitensis challenge was assessed by quantifying bacterial loads in the spleens of infected mice. Mice immunized with Lpp-OmpA-BtaE + FA displayed a significantly lower bacterial burden compared to both the Lpp-OmpA-BtaE and PBS control groups (Fig. 5). Lpp-OmpA-BtaE + FA immunization resulted in the most evident reduction in bacterial counts (up to a 102-fold decrease; P < 0.0001), highlighting its superior protective capabilities. Even Lpp-OmpA-BtaE immunization alone led to a nearly 10-fold reduction in bacterial burden compared to PBS controls (P < 0.002), indicating partial protection.

Lpp-OmpA-BtaE immunization protected mice against B. melitensis infection. Thirty days after the final immunization, mice were challenged intraperitoneally with B. melitensis, and bacterial loads in the spleen were assessed 14 days post-infection. The data represent the mean ± SD of 5 mice per group. **P < 0.004***P < 0.002 and ****P < 0.0001.
Discussion
In this study, the enhanced immunogenic response and protective efficacy of the BtaE antigen against B. melitensishas been demonstrated in a murine model through its incorporation into the Lpp-OmpA-BtaE construct. The integration of computational modeling, epitope prediction, and experimental validation has facilitated the recognition of surface-exposed loops BtaE spanning positions 27–41, 67–83, and 109–209 within the Lpp-OmpA-BtaE fusion protein as potential B-cell epitopes26,27. Secondary structure analysis and 3D modeling have affirmed the linear and discontinuous characteristics of the anticipated BtaE epitopes within the outer loops of the Lpp-OmpA-BtaE fusion protein, thus validating their potential as B-cell epitopes27,28,29. This study’s identification of high-affinity binding peptides across diverse MHC-I and MHC-II alleles strengthens the argument for the Lpp-OmpA-BtaE fusion protein as a potential T-cell vaccine candidate against B. melitensis. These results highlight the Lpp-OmpA-BtaE fusion protein’s ability to bind multiple HLA alleles, suggesting broad population coverage and the potential to induce robust and diverse immune responses30. This broad HLA allele coverage is critical for the development of a vaccine capable of eliciting strong T-cell-mediated immunity against B. melitensis, ensuring efficacy across diverse human populations30,31. Furthermore, the potential for the fusion protein to highlight the distinct roles of MHC class I and II in presenting BtaE antigens to CD8+ and CD4+ T cells, respectively, underscores its capacity to generate comprehensive protective immunity against B. melitensis infection11,32. The MHC class I pathway facilitates the activation of CD8+ cytotoxic T cells, essential for targeting and eliminating infected cells, while the MHC class II pathway aids in the activation of CD4+ helper T cells, crucial for orchestrating a robust immune response11,30,31. Moreover, structural modeling has revealed a notable 99% similarity to a known cell surface protein identified in B. melitensis. The observed similarity implies a potential evolutionary or functional association between the investigated protein and the B. melitensis protein, indicative of conserved structural motifs or functional domains with relevance to immunological contexts, notably in vaccine development14,17,18.
TheLpp-OmpA-BtaE as a fusion protein antigen, facilitates the outer membrane tethering of B. melitensis BtaE33,34,35. Immunization with Lpp-OmpA-BtaE + FA induces a robust humoral immune response characterized by elevated serum IgG, IgG1, and IgG2a titers35,36,37. The analysis of antigen-specific IgG subclasses provides critical insights into immune polarization. Notably, analysis of IgG subclasses reveals a prevalence of IgG2a over IgG1, indicative of a skewed Th1-biased response, is correlated with macrophage activation, improved antigen presentation, and facilitation of bacterial clearance, thereby aiding in the establishment of enduring protective immunity against B. melitensis infection38,39,40. Conversely, IgG1, primarily associated with Th2-driven responses, supports humoral immunity through enhanced antibody production and complement activation41. The concurrent elevation of IgG1 and IgG2a reflects an immune response, where the Lpp-OmpA-BtaE fusion protein, combined with Freund’s adjuvant, stimulates both neutralizing antibody production and cellular immunitykey factors in defense againstB. melitensis42. The use of CFA for priming, followed by IFA booster doses, likely sustains IgG2a responses, as CFA enhances antigen-presenting cell activation and IFN-γ secretion, reinforcing Th1-driven immunity43. The dual induction of IgG subclasses highlights the potential of Lpp-OmpA-BtaE to generate a strong and long-lasting immune response, effectively targeting B. melitensis41,42. Differences in IgG subclass levels relative to total IgG may be attributed to variations in assay sensitivity and antibody kinetics post-immunization. ELISA detection sensitivity, particularly for IgG subclasses, may account for the observed higher values, as subclass-specific antibodies often exhibit stronger binding affinities, leading to enhanced signal intensities44. Additionally, IgG subclass production follows distinct kinetics, with IgG1 and IgG2a peaking at later time points. By day 42, total IgG levels converged with those of the subclasses, reflecting immune maturation and stabilization. The Lpp-OmpA system acts as a molecular scaffold, facilitating the fusion of the BtaE antigen onto the outer membrane of E. coli. The physical coupling of the BtaE antigen to the outer membrane surface through the Lpp-OmpA23,24. The Lpp moiety directs the fusion protein to the outer membrane of E. coli cells, while the OmpA segment promotes surface exposure. The physical coupling of the BtaE antigen to the outer membrane surface through the Lpp-OmpA is pivotal for eliciting robust antibody responses23. The Lpp-OmpA surface displays enhancing accessibility and promoting B-cell recognition of the BtaE antigen on the outer membrane of E. coli cells, consequently eliciting a robust humoral immune response characterized by increased production of IgG, IgG1, and IgG2a antibodies, thereby positioning it as a valuable strategy in the development of an effective Lpp-OmpA-based vaccine against B. melitensis infections45.
The detection of key cytokines, including IFN-γ, IL-12, IL-4, and IL-10, in splenocyte culture supernatants following Lpp-OmpA-BtaE stimulation provides crucial insights into the immune response elicited by this fusion protein. IFN-γ and IL-12 are hallmark Th1 cytokines essential for cell-mediated immunity, a crucial defense against B. melitensis46. IFN-γ promotes macrophage activation, enhances antigen presentation, and stimulates the production of IgG2a, facilitating bacterial clearance47. Additionally, it upregulates MHC class II expression, strengthening adaptive immunity against B. melitensis upon re-exposure48. IL-12, primarily secreted by DCs and macrophages, acts as a Th1-polarizing cytokine by promoting IFN-γ production and activating cytotoxic T cells, both of which are crucial for controllingB. melitensis infections49. It also bridges innate and adaptive immunity by stimulating natural killer cells and enhancing antigen presentation, reinforcing the immune response50.Conversely, IL-4 and IL-10 are Th2-associated cytokines involved in humoral immunity and immune regulation51. IL-4 plays a pivotal role in B-cell proliferation, IgG1 class switching, and antibody-mediated responses, suggesting that Lpp-OmpA-BtaE also stimulates B-cell activation for long-term immunity52. IL-10 functions as an immunoregulatory cytokine, mitigating excessive inflammation while preserving protective immunity53. Elevated IL-10 levels contribute to immunological memory, crucial for long-term protection and recall responses54. The simultaneous detection of Th1 and Th2 cytokines highlights the ability of Lpp-OmpA-BtaE to induce an immune response, integrating both cellular and humoral immunitya key attribute of effective vaccine candidates against B.melitensis55,56.
The observed Th1 bias, characterized by increased IFN-γ and IL-12 levels, underscores the potential of Lpp-OmpA-BtaE to enhance protection against intracellular Brucella species while maintaining immune regulation through IL-4 and IL-1057,58. The cytokine profile further confirms the strong immunogenicity of Lpp-OmpA-BtaE, demonstrating its capacity to prime and activate antigen-specific T and B cells in splenocyte cultures.It seems the Lpp-OmpA surface display system facilitates the efficient display and presentation of the BtaE antigen to APCs to induce stronger activation and proliferation of B and T lymphocytes59.DCs, as professional APCs, play a central role in orchestrating immune responses by processing and presenting antigens through MHC class I and II pathways60. Lpp-OmpA-BtaE + FA may enhance DC activation, leading to upregulation of co-stimulatory molecules such as CD80 and CD86, which are critical for optimal T-cell priming60. This interaction further drives naive CD4+ T-cell differentiation into Th1 effectors that secrete IFN-γ, reinforcing the cell-mediated immune response required for intracellular bacterial clearance61. Future studies employing flow cytometry to assess DC maturation markers and antigen uptake efficiency will provide deeper mechanistic insights into the immunostimulatory properties of the Lpp-OmpA system. Additionally, evaluating cytokine receptor expression on T and B cells post-immunization will clarify interleukin-mediated regulation of adaptive immunity. To accurately assess vaccine-induced responses, quantifying interleukin levels at multiple time points will offer a comprehensive understanding of immune balance. Strategic adjuvant selection and antigen engineering can further optimize cytokine responses, enhancing vaccine efficacy and ensuring long-term protection against brucellosis.Future investigations, particularly in vivo challenge studies, are crucial for validating the protective efficacy of this vaccine candidate by examining cytokine kinetics, antigen-specific T-cell responses, and bacterial clearance. Longitudinal studies tracking cytokine dynamics over time will also provide valuable insights into immune memory formation and the durability of protection conferred by Lpp-OmpA-BtaE + FA immunization. Overall, the cytokine profile elicited by Lpp-OmpA-BtaE + FA suggests a robust and long-lasting immune response, reinforcing its potential as a viable vaccine candidate against B. melitensis.
Several studies have investigated the immunogenic potential of recombinant Brucella antigens, reporting varying degrees of protection. For example, vaccination with recombinant Omp16 elicited a strong Th1 response, characterized by elevated IFN-γ production, and conferred significant protection against B. abortus infection in murine models62. Similarly, studies on the Brucella ribosomal protein L7/L12 have demonstrated robust antigen-specific IFN-γ responses, indicative of effective T-cell activation63,64. However, Omp31, despite eliciting strong humoral responses, has demonstrated a relatively weak Th1 response, underscoring the need for antigen engineering strategies to enhance vaccine efficacy65,66,67,68,69. Our study builds on these findings by integrating the Lpp-OmpA and BtaE sequences into a single fusion construct, which exhibits superior IFN-γ and IL-12 induction compared to previously reported single-antigen vaccines. This antigen fusion strategy enhances the protective efficacy of Brucella subunit vaccines by simultaneously improving Th1-skewed immune responses and balancing pro- and anti-inflammatory cytokine profiles. While Omp19 has been identified as a promising antigen, our results suggest that the Lpp-OmpA-BtaE fusion construct not only enhances IFN-γ production but also modulates IL-10 levels, potentially mitigating excessive inflammation70,71. The observed increase in antigen-specific T-cell responses and cytokine secretion aligns with findings from other Brucella antigens, further validating the immunogenic potential of our construct72.Beyond Brucella vaccine development, the Lpp-OmpA system has been widely employed to enhance antigen presentation and immunogenicity. In a study on Vibrio anguillarum, a modified Lpp-OmpA-based outer membrane anchoring system successfully displayed green fluorescent protein (GFP) on E. coli surfaces, demonstrating the versatility of this approach for vaccine applications73. Similarly, Lpp-OmpA fusion strategies have been utilized in the development of OMV-based vaccines, such as decorating E. coli K-12 OMVs with the receptor-binding domain (RBD) of the SARS-CoV-2 spike protein, which elicited specific immune responses in preclinical models25. A recent study further demonstrated the efficacy of Lpp-OmpA-based OMVs in a COVID-19 vaccine, leveraging an adjuvanted delivery system to enhance immune responses74. These studies collectively highlight the advantages of the Lpp-OmpA system in vaccine design, reinforcing its applicability in developing antigen display platforms across multiple pathogens. Our findings further extend this approach by demonstrating that Lpp-OmpA fusion with Brucella antigens can generate a more comprehensive immune response, addressing the limitations observed in single-antigen vaccine formulations. While our findings demonstrate the immunogenic potential of the Lpp-OmpA-BtaE fusion protein, additional challenge experiments and long-term immunological assessments are necessary to fully establish its protective efficacy relative to existing recombinant vaccine candidates. The potential of Lpp-OmpA-based antigen presentation in combination with OMVs also warrants further investigation as a novel strategy for Brucella vaccine development.
Immunization with Lpp-OmpA-BtaE + FA substantially augmented the populations of both CD8+ and CD4+ T cells, indicative of robust cellular immunity, essential for an effective vaccine targeting the intracellular B. melitensis pathogen32,75,76. Lpp-OmpA-BtaE + FA elicits the generation of CD8+ T cells, which confer direct cell-mediated immunity by identifying and eliminating Brucella-infected host cells77,78. Alongside, the expanded CD4+ T cell population activates CD8+ T cells and enhances the priming of specific antibody responses, critical for opsonization and complement activation against the pathogen32,37. This underscores the vaccine’s potential to confer protective immunity against Brucella infections. The Lpp-OmpA-BtaEantigen achieves augmentation of CD8+ and CD4+ T cell populations through surface presentation of the BtaE antigen, thereby facilitating enhanced accessibility and presentation of APCs such as dendritic cells59,79. Surface display BtaE in the Lpp-OmpA display system likely enhances the immunogenicity of the BtaE antigen through the adjuvant’s capacity to stimulate innate immune cells and provide co-stimulatory signals, thereby activating the proliferation of antigen-specific CD8+ and CD4+ T cells against B. melitensis59,75,79. Elevated production of IFN-γ, IL-12 and IL-4 by Lpp-OmpA-BtaE + FA immune splenocytes signifies activation of CD8+ cytotoxic and CD4+ helper T cells32,75,76. The increased levels of IL-10 and the expanded T cell populations induced by Lpp-OmpA-BtaE immunization can be attributed to the differentiation of CD8+ and CD4+ T cells into long-lived memory subsets, facilitating rapid recall responses upon subsequent B. melitensis exposure80,81. Also, the expansion of Brucella-specific T cell populations holds promise for therapeutic interventions in chronic or persistent infections32,75,76. By enabling efficient antigen presentation, harnessing adjuvant effects, eliciting Th1 and Th2 responses, and fostering immunological memory, the Lpp-OmpA-BtaE vaccine is particularly adept at priming and expanding antigen-specific CD8+ cytotoxic T cells and CD4+ helper T cells, which are pivotal for generating protective cell-mediated and humoral immunity against Brucella infections32,76,81.
In this study, we observed distinct cytokine profiles across different phases of immune activation, particularly for IFN-γ, IL-12, IL-4, and IL-10. The induction of a Th1-skewed response, characterized by elevated IFN-γ and IL-12, is generally associated with enhanced protection against Brucella infection. However, excessive or prolonged cytokine activation may lead to immunopathological consequences, including chronic inflammation, tissue damage, and immune exhaustion, potentially compromising vaccine safety and efficacy82. A critical concern is the overproduction of IFN-γ, which plays a key role in intracellular bacterial clearance but, when persistently elevated, has been implicated in tissue damage and immunopathology82. Prolonged IFN-γ exposure can result in macrophage overactivation and excessive production of reactive oxygen species (ROS) and nitric oxide (NO), contributing to oxidative stress and cytotoxicity in host tissues46. This highlights the necessity of achieving a balanced cytokine response to facilitate effective pathogen clearance while mitigating inflammatory damage. Similarly, IL-12, essential for Th1 differentiation, can drive sustained IFN-γ secretion, exacerbating inflammatory responses and increasing the risk of tissue fibrosis and autoimmune pathology83. Long-term assessments of IL-12 upregulation in Brucella vaccination models are warranted to evaluate its impact on immune homeostasis84. Conversely, IL-4 and IL-10, which regulate Th2 and Treg responses, respectively, exhibited variable expression across different phases of immune activation. While IL-10 plays a crucial role in controlling excessive inflammation, prolonged elevation may suppress Th1-mediated immunity, impair bacterial clearance, and contribute to pathogen persistence85,86. Elevated IL-10 levels have been observed in chronic Brucella infections, where they facilitate immune evasion by inhibiting macrophage activation and antigen presentation87. While transient IL-10 expression may help mitigate excessive inflammation, its sustained production could undermine vaccine efficacy.Given these findings, future studies should focus on refining cytokine modulation strategies to optimize immune responses while minimizing potential adverse effects. This may include strategic adjuvant selection, antigen dose optimization, and immunoregulatory checkpoint analysis. Additionally, advanced immunological profiling techniques, such as multiplex cytokine assays and transcriptomic analyses, should be employed to assess the broader impact of cytokine alterations on host immune networks. While the observed cytokine profiles suggest the induction of protective immunity, a deeper understanding of the balance between immune activation and regulation is essential to ensure that vaccine-induced responses are both effective and safe.
Immunization with Lpp-OmpA-BtaE + FA resulted in a remarkable reduction in the bacterial load in the spleens of B. melitensis-infected mice, a pivotal indicator of the vaccine’s potential efficacy in conferring protection against brucellosis. The Lpp-OmpA-BtaEinjection induced elevated serum IgG antibody titers, which contribute to bacterial clearance by promoting opsonophagocytosis by macrophages, activating the complement system, and neutralizing bacterial virulence factors88,89. Furthermore, Lpp-OmpA-BtaE immunization led to significant increases in CD8+ cytotoxic T cells and CD4+ helper T cells, both of which play essential roles in combatting the intracellular B. melitensis pathogen32,75,76. The combined effect of robust antibody responses and potent cell-mediated immunity elicited by Lpp-OmpA-BtaE likely acts synergistically to diminish bacterial load in Brucella-infected mice.
This study evaluated the protective efficacy of the Lpp-OmpA-BtaE fusion protein primarily through quantification of bacterial burden in the spleen, a primary target organ for B. melitensis replication in murine models. However, the absence of bacterial load assessments in other organs (e.g., liver, lymph nodes) represents a limitation in fully characterizing the vaccine’s protective potential. Additionally, histopathological evaluation was not performed, which could have provided insights into immune-mediated tissue responses, including inflammation and immune cell infiltration, complementing the observed reduction in splenic bacterial burden. The focus on splenic colonization as the primary readout of vaccine efficacy was based on its established role as a reliable correlate of protection in murine brucellosis models. Nevertheless, a more comprehensive evaluation, including colony-forming unit quantification in secondary organs and survival analysis over a 30-day post-challenge period, is necessary to delineate the breadth and durability of immune protection. Future studies will address these limitations by assessing bacterial loads in the liver and lymph nodes, conducting survival analyses, and incorporating histopathological assessments to evaluate tissue-specific immune responses. While the Lpp-OmpA-BtaE + FA group demonstrated superior protective efficacy, additional assessmentsincluding long-term protection, bacterial clearance kinetics, and tissue-specific immune responsesare necessary to fully define the durability and robustness of immunity. Future studies should employ challenge models in relevant hosts, such as murine or ruminant species, to evaluate organ-specific bacterial loads, persistence of immunity, and comparative efficacy against existing vaccines. The study also highlights the critical role of adjuvant selection in optimizing vaccine immunogenicity. While Lpp-OmpA-BtaE alone significantly reduced bacterial counts, its combination with Freund’s adjuvant (Lpp-OmpA-BtaE + FA) further enhanced protective efficacy. Investigating alternative adjuvants, such as Toll-like receptor agonists or nanoparticle-based systems, may improve antigen presentation and immune activation, leading to more robust and durable responses. Furthermore, the immunological analysis in this study primarily focused on cytokine profiling. A more detailed assessment of antigen-specific T-cell responses, including memory T-cell activation and polyfunctional T-cell characterization, would provide deeper mechanistic insights into cellular immunity. Advanced techniques such as flow cytometry and single-cell RNA sequencing could further refine our understanding of the immune pathways involved.Expanding the antigenic repertoire to include additional Brucella antigens, such as Omp16 and Omp19 may also enhance vaccine coverage and protective efficacy. Future research should integrate histopathological assessments, optimize vaccine formulations with novel adjuvants, employ diverse challenge models, and conduct longitudinal studies on immune memory and cellular responses. These investigations will be instrumental in the development of a more effective and durable Brucella vaccine.
Comprehensive safety evaluation is integral to vaccine development, particularly for recombinant fusion proteins for immunization. While this study primarily assesses the immunogenicity and protective efficacy of the Lpp-OmpA-BtaE fusion protein, future investigations must rigorously evaluate its systemic toxicity and potential off-target effects. Histopathological analyses of vital organs and hematological assessments can provide critical insights into the biological safety profile of the immunogen. Additionally, the risk of unintended immune responses, including hypersensitivity reactions and autoimmune manifestations, underscores the need for long-term immunological monitoring. Given the potential variability in the persistence of recombinant protein-based vaccines within the host, biodistribution studies are essential to elucidate the fusion protein’s clearance kinetics and tissue retention. These comprehensive safety assessments are fundamental to establishing the translational viability of Lpp-OmpA-BtaE as a vaccine candidate against B. melitensis.
In conclusion, incorporating BtaE into the Lpp-OmpA-BtaE construct augments the immunogenic response and protective efficacy of the BtaE antigen against B. melitensis in a murine model. The observed immunological responses, characterized by elevated serum IgG antibody titers, augmented populations of CD8+ and CD4+ T cells, and cytokine production, indicate that Lpp-OmpA-BtaE + FA immunization elicits both humoral and cellular immunity, synergistically contributing to the reduction of bacterial load in infected mice. These findings necessitate a comprehensive assessment of the Lpp-OmpA-BtaEvaccine’s efficacy against a wide range of Brucellaspecies. Also, our data suggests exploring the application of the Lpp-OmpA-BtaEimmunogenecity in other models of B. melitensis infection. Overall, these results emphasize the potential of the Lpp-OmpA-BtaE antigen as a promising strategy against brucellosis.
Materials and methods
Bacterial strains
Escherichia coli strains DH5α and BL21 (DE3) were used for plasmid construct propagation and recombinant protein overexpression, respectively. Both B. melitensis 16 M and E. coli strains were kindly provided by the Department of Bacteriology, Pasteur Institute of Iran (Tehran, Iran). Brucella melitensis 16 M ATCC 23,456 was applied for intraperitoneal infection of mice.
Ethics statement
The study is reported in accordance with ARRIVE guidelines.All methods were performed in accordance with the relevant guidelines and regulations.All animal experimental protocols were approved by the Research Ethics Committees of the Islamic Azad University Science and Research Branch (approval number: IR.IAU.SRB.REC.1401.343). Female BALB/C mice (6–8 weeks old) were obtained from the Pasteur Institute of Iran (Tehran, Iran). The mice were subsequently divided into control and experimental groups, each containing 12 animals. For sample size analysis, 80% power, α = 0.05 determined the sample size for each group for subsequent analyses, including serum antibody titers, lymphocyte proliferation assays, cytokine analysis, and bacterial load determination. For the in vivo experiments, anesthesia was administered to mice using a ketamine-xylazine combination. Euthanasia was subsequently conducted by subjecting the mice to carbon dioxide (CO₂) inhalation from a regulated pressurized tank.
Bioinformatics analysis of Lpp-OmpA-BtaE fusion protein
The amino acid sequences of the BtaE orthologue from B. melitensis (BMEI1873) (GenBank Accession No: AAL53054.1), OmpA (GenBank Accession No: NP_415477.1), and Lpp (Nucleotide Accession No: NC_000913.3) of E. coli K12 MG1655 were retrieved from the National Center for Biotechnology Information (NCBI). The PRED-TMBB tool (RRID: SCR_006190) (http://bioinformatics.biol.uoa.gr/PRED-TMBB/input.jsp) was used to predict surface-exposed regions and identify the protein topology. SignalP 5.1 (RRID: SCR_015644) (https://services.healthtech.dtu.dk/services/SignalP-5.0/) was employed to predict the probable signal peptide within the sequence. Linear B-cell epitopes were predicted and characterized using ABCpred(http://webs.iiitd.edu.in/raghava/abcpred/), BepiPred-1.0 (http://tools.iedb.org/bcell/), BepiPred-2.0 (RRID: SCR_018499)(https://services.healthtech.dtu.dk/service. php? BepiPred-2.0) and LBtope(http://crdd.osdd.net/raghava/lbtope/). Protein secondary structure was predicted using SYMPRED (http://www.ibi.vu.nl/programs/sympredwww/), PSIPRED 4.0 (RRID: SCR_010246) (http://bioinf.cs.ucl.ac.uk/psipred) and SSPro (https://download.igb.uci.edu/sspro4.html).ElliPro was utilized to identify linear and discontinuous epitopes based on the 3D structure of the protein antigens. Immune epitope database (IEDB) tools (RRID: SCR_006604) including Emini Surface Accessibility Scale, Karplus & Schulz Flexibility Scale, Parker Hydrophilicity Prediction and Chou and Fasman Beta-Turn Prediction were used to predict and analyze epitopes. Sequence homology was evaluated using PSI-BLAST against the Protein Data Bank (PDB). NetMHCpan-4.1 (RRID: SCR_018182) (http://tools.iedb.org/mhci/) and NetMHCIIpan-4.1 (http://tools.iedb.org/mhcii/),used to predict antigen binding scores to major histocompatibility complex class I and II molecules (MHC class I and MHC class II), respectively. The 3D structures of sequences were predicted using SWISS-MODEL (RRID: SCR_018123) (https://swissmodel.expasy.org), I-TASSER (RRID: SCR_014627)(https://zhanglab.ccmb.med.umich.edu/I-TASSER/) and Robetta (RRID: SCR_018805)(http://robetta.bakerlab.org/) servers. Predicted structures underwent refinement and validation using 3Drefine (RRID: SCR_021883)(https://3drefine.mu.hekademeia.org/index.html), QMEAN (https://swissmodel.expasy.org/qmean/) and RAMPAGE (RRID: SCR_017590) (https://www.ebi.ac.uk/thornton-srv/databases/pdbsum/) tools.
Preparation of recombinant Lpp-OmpA-BtaE fusion protein
The amino acid sequences of fusion protein encoding the Lpp (2–9), OmpA (46–159) and BtaE (16–363) were optimized, synthesized and cloned into the pET26b vector between NdeI and EcoRI by Biomatik Company (Ontario, Canada). The recombinant plasmid was then verified using PCR, restriction enzyme digestion, and DNA sequencing. For protein expression, IPTG (Sigma, USA) at a final concentration of 1 mM was added to the bacterial cultures to induce Lpp-OmpA-BtaE fusion protein expression34. After 4 h of induction, the bacterial cultures were centrifuged at 11,000 g for 30 min at 25 °C to pellet the cells. The cell pellets were lysed in buffer B (100 mM NaH2PO4, 10 mM Tris-HCl, and 8 M urea; pH 8.0) andthen incubated with Ni-NTA agarose resin (QIAGEN, Germany) on a rotary shaker at room temperature (RT) for 1 h. The resin was washed with buffer C (100 mM NaH2PO4, 10 mM Tris-HCl, and 8 M urea; pH 6.3). Urea was gradually removed using washing buffers with decreasing urea concentrations (8, 6, 4, 2, 1, and 0 M). The purified Lpp-OmpA-BtaE fusion protein was finally eluted with buffer E (50 mM NaH2PO4, 20 mM Tris-HCl, and 250 mM imidazole; pH 4.5). To remove imidazole, the eluted protein solution was dialyzed against phosphate-buffered saline (PBS, pH 7.4) overnight. The level of induced protein was determined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) with 12.5% (w/v) polyacrylamide gels followed by Coomassie Brilliant Blue G-250 staining (Bio-Rad, Hercules, CA, USA). To verify the identity and integrity of the purified Lpp-OmpA-BtaE fusion protein, Western blot analysis was performed. Proteins were separated by SDS-PAGE and transferred onto a polyvinylidene difluoride (PVDF) membrane (Bio-Rad) using a semi-dry blotting system. Membranes were blocked with 5% (w/v) non-fat dry milk in Tris-buffered saline with 0.1% Tween-20 (TBST) for 1 h at room temperature to minimize non-specific antibody binding. The membrane was then incubated with a horseradish peroxidase (HRP)-conjugated mouse monoclonal anti-polyhistidine antibody (Sigma-Aldrich) at a 1:2000 dilution in TBST. Immunoreactive bands were detected using 3,3′-diaminobenzidine (DAB) substrate (Sigma-Aldrich) and visualized with a high-resolution digital imaging system. The presence of a distinct band at the expected molecular weight confirmed successful expression and purification of the Lpp-OmpA-BtaE fusion protein. Endotoxin levels in the purified protein were determined using the PYROGENTTM Gel Clot LAL Assay (Charles River Laboratories) and found to be below 10 EU/mL.
Mouse immunizations and B. melitensischallenges
Female BALB/c mice were divided into three groups (n = 12/group). To evaluate the immunogenicity of the Lpp-OmpA-BtaE fusion protein, two immunization regimens were employed. One group received an intraperitoneal (i.p.) injection of 30 µgLpp-OmpA-BtaE in complete Freund’s adjuvant (CFA) on day 0, followed by two booster immunizations with 30 µg of the recombinant protein in incomplete Freund’s adjuvant (IFA) on days 14 and 28. Another group received 30 µg of Lpp-OmpA-BtaE in phosphate-buffered saline (PBS) on day 0, followed by two booster immunizations with 30 µg of the protein in PBS on days 14 and 28. The control group received sterile PBS on the same schedule. Serum samples were collected from the orbital sinus blood on days 14, 28, and 42 to analyze antigen-specific antibody responses.Thirty days after the final immunization, all mice were challenged with B. melitensis 16 M as described previously88. Briefly, mice were infected intraperitoneally with a dose of 5 × 10⁵ colony-forming units (CFU) of B. melitensis 16 M. Fourteen days after the challenge, spleens from experimental mice were aseptically excised, homogenized, serially diluted, and cultured on Tryptic Soy Agar (TSA) plates to quantify bacterial colonies following five days of incubation at 37 °C.
Assessmentof antibody responses
To evaluate the humoral immune response elicited by Lpp-OmpA-BtaE immunization, antigen-specific IgG titers were measured using an enzyme-linked immunosorbent assay (ELISA). Briefly, microtiter plates were coated with 100 µL of Lpp-OmpA-BtaE (10 µg/mL) in carbonate-bicarbonate buffer overnight at 4 °C. The plates were then washed with PBS-Tween (PBS-T) and blocked with PBS containing 3% bovine serum albumin (BSA). Subsequently, serially diluted mouse sera were added to the wells and incubated for 2 h at RT. Following three washes with PBS-T, HRP-conjugated goat anti-mouse IgG secondary antibody (diluted 1:10,000 in PBS-T) was added and incubated for 1 h at RT. After additional washes with PBS-T, the TMB substrate (100 µL/well) was added, and the reaction was stopped with 2 N H2SO4, and the absorbance was measured at 450 nm (OD450). The Serum IgG1 and IgG2a subtype concentrations were determined similarly using specific HRP-conjugated secondary antibodies against mouse IgG1 or IgG2a (BioLegend, USA). The endpointtiter was defined as the highest dilution at which the OD450 value exceeded the background by at least 0.1. Background wells contained PBS instead of serum samples90.
Splenocyte cytokine secretion assay
One month following the final immunization, spleens of experimental mice were harvested, homogenized, and suspended in a complete RPMI 1640 medium containing 2% FBS. Red blood cells (RBCs) were lysed using an established RBC lysis buffer (BioLegend, USA). Then, cells were resuspended in complete RPMI 1640 at a density of 1 × 10⁶ cells/mL and plated in 24-well plates (Nunc, USA). To assess antigen-specific cytokine production, splenocytes were stimulated with 10 µg/mL of Lpp-OmpA-BtaE fusion protein. Phytohemagglutinin-A (PHA; 10 µM; Gibco, UK) served as a positive control for T cell activation, while unstimulated cells were included as a negative control. Following a 72-hour incubation period, cell culture supernatants were collected and analyzed for the presence of interferon-γ (IFN-γ), interleukin-12 (IL-12), interleukin-10 (IL-10), and interleukin-4 (IL-4) using specific ELISA kits (PeproTech, USA) according to the manufacturer’s instructions.
Assessment of T-cell responses
To determine the T cell subpopulations induced by Lpp-OmpA-BtaE immunization, stimulated splenocytes were washed with fluorescence-activated cell sorting (FACS) buffer (PBS containing 2% FBS) and stained with fluorescence-conjugated antibodies. Cells were incubated with Cy-chrome-conjugated anti-CD8+ (clone 53 − 6.7; BioLegend, USA) and fluorescein isothiocyanate (FITC)-conjugated anti-CD4+ (clone GK1.5; BioLegend, USA) monoclonal antibodies specific for cytotoxic T lymphocytes (CTLs) and T helper cells, respectively. After additional washes with FACS buffer, fluorescence intensity was measured using a BD Fortessa flow cytometer (BD Biosciences, USA) equipped with BD FACSDiva software. Flow cytometry data were analyzed using FlowJo software (TreeStar, USA) to quantify the percentages of CD8+ and CD4+ T cell populations within the stimulated splenocyte cultures.
Statistical analyses were performed using GraphPad Prism 8 (GraphPad Software, USA). One-way analysis of variance (ANOVA) was employed to assess differences between groups for all measured parameters (antibody titers, cytokine levels, and T-cell subpopulations). Dunnett’s post hoc test was used for multiple comparisons between the immunized groups and the control group. Results are expressed as the mean ± standard deviation (SD). A p-value of less than 0.05 (P < 0.05) was considered statistically significant.
Data availability
Upon a reasonable request, the corresponding author will provide access to the datasets generated or analyzed during the current study.
References
-
Czibener, C. & Ugalde, J. E. Identification of a unique gene cluster of Brucella spp. That mediates adhesion to host cells. Microbes Infect. 14, 79–85. https://doi.org/10.1016/j.micinf.2011.08.012 (2012).
-
Huleatt, J. W. et al. Potent immunogenicity and efficacy of a universal influenza vaccine candidate comprising a recombinant fusion protein linking influenza M2e to the TLR5 ligand flagellin. Vaccine26, 201–214, (2008). https://doi.org/10.1016/j.vaccine.2007.10.062
-
Bennett, K. M. et al. Hybrid Flagellin as a T cell independent vaccine scaffold. BMC Biotechnol. 15, 015–0194. (2015).
-
Franco, M. P., Mulder, M., Gilman, R. H. & Smits, H. L. Human brucellosis. Lancet Infect. Dis. 7, 775–786. https://doi.org/10.1016/S1473-3099(07)70286-4 (2007).
-
Pappas, G., Papadimitriou, P., Akritidis, N., Christou, L. & Tsianos, E. V. The new global map of human brucellosis. Lancet Infect. Dis. 6, 91–99. https://doi.org/10.1016/S1473-3099(06)70382-6 (2006).
-
Moreno, E., Blasco, J. M. & Moriyón, I. Facing the Human and Animal Brucellosis Conundrums: The Forgotten Lessons. Microorganisms10 (2022).
-
Nikbakht, M. & Pakbin, B. Nikbakht brujeni, G. Evaluation of a new lymphocyte proliferation assay based on Cyclic voltammetry; an alternative method. Sci. Rep. 9 https://doi.org/10.1038/s41598-019-41171-8 (2019).
-
Avila-Calderon, E. D., Lopez-Merino, A., Sriranganathan, N., Boyle, S. M. & Contreras-Rodriguez A. A history of the development of Brucella vaccines. Biomed. Res. Int. 743509, (2013). https://doi.org/10.1155/2013/743509 (2013).
-
Roca, I., Espinal, Vila-Farres, X. & Vila, J. The Acinetobacter baumannii oxymoron: commensal hospital dweller turned Pan-Drug-Resistant menace. Front. Microbiol. 3 (148). https://doi.org/10.3389/fmicb.2012.00148 (2012).
-
Elbehiry, A. et al. The development of diagnostic and vaccine strategies for early detection and control of human brucellosis, particularly in endemic areas. Vaccines (Basel. 11 https://doi.org/10.3390/vaccines11030654 (2023).
-
Heidary, M. et al. Evaluation of brucellosis vaccines: A comprehensive review. Front. Vet. Sci. 9 https://doi.org/10.3389/fvets.2022.925773 (2022).
-
Darbandi, A. et al. Evaluation of immune responses to Brucella vaccines in mouse models: A systematic review. Front. Vet. Sci. 9 https://doi.org/10.3389/fvets.2022.903890 (2022).
-
Linke, D., Riess, T., Autenrieth, I. B., Lupas, A. & Kempf, V. A. Trimeric autotransporter adhesins: variable structure, common function. Trends Microbiol. 14, 264–270. https://doi.org/10.1016/j.tim.2006.04.005 (2006).
-
Lyskowski, A., Leo, J. C. & Goldman, A. Structure and biology of trimeric autotransporter adhesins. Adv. Exp. Med. Biol. 715, 143–158. https://doi.org/10.1007/978-94-007-0940-9_9 (2011).
-
Cotter, S. E., Surana, N. K. & St Geme, J. W. 3 Trimeric autotransporters: a distinct subfamily of autotransporter proteins. Trends Microbiol. 13, 199–205. https://doi.org/10.1016/j.tim.2005.03.004 (2005).
-
Kline, K. A., Falker, S., Dahlberg, S., Normark, S. & Henriques-Normark, B. Bacterial adhesins in host-microbe interactions. Cell. Host Microb.. 5, 580–592. https://doi.org/10.1016/j.chom.2009.05.011 (2009).
-
Ruiz-Ranwez, V. et al. BtaE, an adhesin that belongs to the trimeric autotransporter family, is required for full virulence and defines a specific adhesive pole of Brucella suis. Infect. Immun. 81, 996–1007. https://doi.org/10.1128/IAI.01241-12 (2013).
-
Munoz Gonzalez, F. et al. The BtaF Adhesin Is Necessary for Full Virulence During Respiratory Infection by Brucella suis and Is a Novel Immunogen for Nasal Vaccination Against Brucella Infection. Front Immunol.10, 1775, (2019). https://doi.org/10.3389/fimmu.2019.01775
-
Georgiou, G. et al. Display of heterologous proteins on the surface of microorganisms: from the screening of combinatorial libraries to live Recombinant vaccines. Nat. Biotechnol. 15, 29–34. https://doi.org/10.1038/nbt0197-29 (1997).
-
Lower, B. H., Yongsunthon, R., Vellano, F. P., Lower, S. K. & 3rd & Simultaneous force and fluorescence measurements of a protein that forms a bond between a living bacterium and a solid surface. J. Bacteriol. 187, 2127–2137. https://doi.org/10.1128/jb.187.6.2127-2137.2005 (2005).
-
Francisco, J. A., Earhart, C. F. & Georgiou, G. Transport and anchoring of beta-lactamase to the external surface of Escherichia coli. Proc. Natl. Acad. Sci. U S. A89, 2713–2717. https://doi.org/10.1073/pnas.89.7.2713 (1992).
-
Huang, G. L., Anderson, T. D. & Clubb, R. T. Engineering microbial surfaces to degrade lignocellulosic biomass. Bioengineered 5, 96–106. https://doi.org/10.4161/bioe.27461 (2014).
-
Jeiranikhameneh, M., Razavi, M. R., Irani, S., Siadat, S. D. & Oloomi, M. Designing novel construction for cell surface display of protein E on Escherichia coli using non-classical pathway based on Lpp-OmpA. AMB Express.. 7 https://doi.org/10.1186/s13568-017-0350-0 (2017).
-
Park, K. S. et al. Detoxified synthetic bacterial membrane vesicles as a vaccine platform against bacteria and SARS-CoV-2. J. Nanobiotechnol. 21 https://doi.org/10.1186/s12951-023-01928-w (2023).
-
Thapa, H. B., Muller, A. M., Camilli, A. & Schild, S. An intranasal vaccine based on outer membrane vesicles against SARS-CoV-2. Front. Microbiol. 12 (752739). https://doi.org/10.3389/fmicb.2021.752739 (2021).
-
Nicchi, S. et al. Decorating the surface of Escherichia coli with bacterial lipoproteins: a comparative analysis of different display systems. Microb. Cell. Fact. 20 https://doi.org/10.1186/s12934-021-01528-z (2021).
-
Thibau, A. et al. Immunogenicity of trimeric autotransporter adhesins and their potential as vaccine targets. Med. Microbiol. Immunol. 209, 243–263. https://doi.org/10.1007/s00430-019-00649-y (2020).
-
Bhattacharya, D., Nowotny, J., Cao, R. & Cheng, J. 3Drefine: an interactive web server for efficient protein structure refinement. Nucl. Acids Res. 44 https://doi.org/10.1093/nar/gkw336 (2016).
-
Kim, D. E., Chivian, D. & Baker, D. Protein structure prediction and analysis using the Robetta server. Nucl. Acids Res. 32 https://doi.org/10.1093/nar/gkh468 (2004).
-
Durward, M. A., Harms, J., Magnani, D. M., Eskra, L. & Splitter, G. A. Discordant Brucella melitensis antigens yield cognate CD8 + T cells in vivo. Infect. Immun. 78, 168–176. https://doi.org/10.1128/IAI.00994-09 (2010).
-
Cannella, A. P. et al. Brucella melitensis T cell epitope recognition in humans with brucellosis in Peru. Infect. Immun. 82, 124–131. https://doi.org/10.1128/IAI.00796-13 (2014).
-
Hanot Mambres, D. et al. Identification of immune effectors essential to the control of primary and secondary intranasal infection with Brucella melitensis in mice. J. Immunol. 196, 3780–3793. https://doi.org/10.4049/jimmunol.1502265 (2016).
-
Lieberman, L. A. Outer membrane vesicles: A bacterial-derived vaccination system. Front. Microbiol. 13 https://doi.org/10.3389/fmicb.2022.1029146 (2022).
-
Micoli, F., Adamo, R. & Nakakana, U. Outer membrane vesicle vaccine platforms. BioDrugs 38, 47–59. https://doi.org/10.1007/s40259-023-00627-0 (2024).
-
Weyant, K. B. et al. A modular vaccine platform enabled by decoration of bacterial outer membrane vesicles with biotinylated antigens. Nat. Commun. 14 https://doi.org/10.1038/s41467-023-36101-2 (2023).
-
Konig, E. et al. Multi-Antigen outer membrane vesicle engineering to develop polyvalent vaccines: the Staphylococcus aureus case. Front. Immunol. 12 (752168). https://doi.org/10.3389/fimmu.2021.752168 (2021).
-
Vitry, M. A. et al. Humoral immunity and CD4 + Th1 cells are both necessary for a fully protective immune response upon secondary infection with Brucella melitensis. J. Immunol. 192, 3740–3752. https://doi.org/10.4049/jimmunol.1302561 (2014).
-
Mailybayeva, A. et al. Evaluation of duration of immunogenicity and protective efficacy of improved influenza viral Vector-Based Brucella abortus vaccine against Brucella melitensis infection in sheep and goats. Front. Vet. Sci. 7 https://doi.org/10.3389/fvets.2020.00058 (2020).
-
Mailybayeva, A. et al. Improved influenza viral vector based Brucella abortus vaccine induces robust B and T-cell responses and protection against Brucella melitensis infection in pregnant sheep and goats. PLoS One. 12, e0186484. https://doi.org/10.1371/journal.pone.0186484 (2017).
-
Sanakkayala, N. et al. Induction of antigen-specific Th1-type immune responses by gamma-irradiated Recombinant Brucella abortus RB51. Clin. Diagn. Lab. Immunol. 12, 1429–1436. https://doi.org/10.1128/CDLI.12.12.1429-1436.2005 (2005).
-
Bretscher, P. A. On the mechanism determining the TH1/TH2 phenotype of an immune response, and its pertinence to strategies for the prevention, and treatment, of certain infectious diseases. Scand. J. Immunol. 79, 361–376. https://doi.org/10.1111/sji.12175 (2014).
-
Skendros, P., Pappas, G. & Boura, P. Cell-mediated immunity in human brucellosis. Microbes Infect. 13, 134–142. https://doi.org/10.1016/j.micinf.2010.10.015 (2011).
-
Billiau, A. & Matthys, P. Interferon-gamma: a historical perspective. Cytokine Growth Factor. Rev. 20, 97–113. https://doi.org/10.1016/j.cytogfr.2009.02.004 (2009).
-
Mestas, J. & Hughes, C. C. Of mice and not men: differences between mouse and human immunology. J. Immunol. 172, 2731–2738. https://doi.org/10.4049/jimmunol.172.5.2731 (2004).
-
Vahed, M. et al. Molecular dynamics simulation and experimental study of the surface-display of SPA protein via Lpp-OmpA system for screening of IgG. AMB Express. 10 (161). https://doi.org/10.1186/s13568-020-01097-1 (2020).
-
Ritchie, J. A., Rupper, A., Cardelli, J. A. & Bellaire, B. H. Host interferon-γ inducible protein contributes to Brucella survival. Front. Cell. Infect. Microbiol. 2 https://doi.org/10.3389/fcimb.2012.00055 (2012).
-
Xu, G. et al. Dynamic Changes of Th1 Cytokines and the Clinical Significance of the IFN-γ/TNF-α Ratio in Acute Brucellosis. Mediators Inflamm. 5869257, (2019). https://doi.org/10.1155/2019/5869257 (2019).
-
Pellegrini, J. M., Gorvel, J. P. & Mémet, S. Immunosuppressive mechanisms in brucellosis in light of chronic bacterial diseases. Microorganisms 10, 103390microorganisms10071260 (2022).
-
Trinchieri, G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 3, 133–146. https://doi.org/10.1038/nri1001 (2003).
-
Watford, W. T., Moriguchi, M., Morinobu, A. & O’Shea, J. J. The biology of IL-12: coordinating innate and adaptive immune responses. Cytokine Growth Factor. Rev. 14, 361–368. https://doi.org/10.1016/s1359-6101(03)00043-1 (2003).
-
Mitchell, R. E. et al. IL-4 enhances IL-10 production in Th1 cells: implications for Th1 and Th2 regulation. Sci. Rep. 7 https://doi.org/10.1038/s41598-017-11803-y (2017).
-
Priyanka, Shringi, B. N., Choudhary, O. P. & Kashyap, S. K. Cytokines in brucellosis: biological rhythm at the interface of innate and adaptive immunity. Biol. Rhythm Res. 52, 1031–1043. https://doi.org/10.1080/09291016.2019.1613794 (2021).
-
Couper, K. N., Blount, D. G. & Riley, E. M. IL-10: the master regulator of immunity to infection. J. Immunol. 180, 5771–5777. https://doi.org/10.4049/jimmunol.180.9.5771 (2008).
-
Stranahan, L. et al. Primary and memory immune responses against rough Brucella canis are less robust compared to smooth B. abortus and B. melitensis following intratracheal infection in mice. Front. Immunol. 13, 959328. https://doi.org/10.3389/fimmu.2022.959328 (2022).
-
Vitry, M. A. et al. Crucial role of gamma interferon-producing CD4 + Th1 cells but dispensable function of CD8 + T cell, B cell, Th2, and Th17 responses in the control of Brucella melitensis infection in mice. Infection Immunity80, 4271–4280, (2012). https://doi.org/10.1128/IAI.00761-12
-
Sathiyaseelan, J. et al. Treatment of Brucella-susceptible mice with IL-12 increases primary and secondary immunity. Cell. Immunol. 243, 1–9. https://doi.org/10.1016/j.cellimm.2006.10.003 (2006).
-
Li, T. et al. Brucella melitensis M5-90Deltabp26 as a potential live vaccine that allows for the distinction between natural infection and immunization. Can. J. Microbiol. 63, 719–729. https://doi.org/10.1139/cjm-2017-0179 (2017).
-
Eze, M. O. et al. Effects of opsonization and gamma interferon on growth of Brucella melitensis 16 M in mouse peritoneal macrophages in vitro. Infection and Immun.68, 257–263, (2000). https://doi.org/10.1128/IAI.68.1.257-263.2000
-
Cheng, K. et al. Bioengineered bacteria-derived outer membrane vesicles as a versatile antigen display platform for tumor vaccination via Plug-and-Display technology. Nat. Commun. 12, 2041. https://doi.org/10.1038/s41467-021-22308-8 (2021).
-
Théry, C. & Amigorena, S. The cell biology of antigen presentation in dendritic cells. Curr. Opin. Immunol. 13, 45–51. https://doi.org/10.1016/s0952-7915(00)00180-1 (2001).
-
Zanna, M. Y. et al. Review of dendritic cells, their role in clinical immunology, and distribution in various animal species. Int. J. Mol. Sci. 22 https://doi.org/10.3390/ijms22158044 (2021).
-
Pasquevich, K. A. et al. Immunization with Recombinant Brucella species outer membrane protein Omp16 or Omp19 in adjuvant induces specific CD4 + and CD8 + T cells as well as systemic and oral protection against Brucella abortus infection. Infect. Immun. 77, 436–445. https://doi.org/10.1128/iai.01151-08 (2009).
-
Zhu, L. et al. Comparison of immune effects between Brucella Recombinant Omp10-Omp28-L7/L12 proteins expressed in eukaryotic and prokaryotic systems. Front. Vet. Sci. 7 https://doi.org/10.3389/fvets.2020.00576 (2020).
-
Oliveira, S. C. & Splitter, G. A. Subcloning and expression of the Brucella abortus L7/L12 ribosomal gene and T-lymphocyte recognition of the Recombinant protein. Infect. Immun. 62, 5201–5204. https://doi.org/10.1128/iai.62.11.5201-5204.1994 (1994).
-
Cassataro, J. et al. Vaccination with the Recombinant Brucella outer membrane protein 31 or a derived 27-amino-acid synthetic peptide elicits a CD4 + T helper 1 response that protects against Brucella melitensis infection. Infect. Immun. 73, 8079–8088. https://doi.org/10.1128/iai.73.12.8079-8088.2005 (2005).
-
Perkins, S. D., Smither, S. J. & Atkins, H. S. Towards a Brucella vaccine for humans. FEMS Microbiol. Rev. 34, 379–394. https://doi.org/10.1111/j.1574-6976.2010.00211.x (2010).
-
Golshani, M. et al. Evaluation of Poly(I:C) and combination of CpG ODN plus Montanide ISA adjuvants to enhance the efficacy of outer membrane vesicles as an acellular vaccine against Brucella melitensis infection in mice. Int. Immunopharmacol. 84, 106573. https://doi.org/10.1016/j.intimp.2020.106573 (2020).
-
Mohammadi, E. & Golchin, M. High protection of mice against Brucella abortus by oral immunization with recombinant probiotic Lactobacillus casei vector vaccine, expressing the outer membrane protein OMP19 of Brucella species. Compar. Immunol. Microbiol. Infect. Dis.70, 101470 (2020).
-
Naseer, A., Mo, S., Olsen, S. C. & McCluskey, B. Brucella melitensis vaccines: A systematic review. Agriculture 13, 2137 (2023).
-
Darriba, M. L., Cerutti, M. L., Bruno, L., Cassataro, J. & Pasquevich, K. A. Stability studies of the vaccine adjuvant U-Omp19. J. Pharm. Sci. 110, 707–718. https://doi.org/10.1016/j.xphs.2020.10.011 (2021).
-
Tadepalli, G., Singh, A. K., Balakrishna, K., Murali, H. S. & Batra, H. V. Immunogenicity and protective efficacy of Brucella abortus Recombinant protein cocktail (rOmp19 + rP39) against B. abortus 544 and B. melitensis 16 M infection in murine model. Mol. Immunol. 71, 34–41. https://doi.org/10.1016/j.molimm.2016.01.001 (2016).
-
Lalsiamthara, J. & Lee, J. H. Development and trial of vaccines against Brucella. J. Vet. Sci. 18, 281–290. https://doi.org/10.4142/jvs.2017.18.S1.281 (2017).
-
Yang, Z., Liu, Q., Wang, Q. & Zhang, Y. Novel bacterial surface display systems based on outer membrane anchoring elements from the marine bacterium Vibrio anguillarum. Appl. Environ. Microbiol. 74, 4359–4365. https://doi.org/10.1128/aem.02499-07 (2008).
-
Yılmaz Çolak, Ç. Bacterial membrane vesicles as a novel vaccine platform against SARS-CoV-2. Curr. Microbiol. 81 https://doi.org/10.1007/s00284-024-03846-y (2024).
-
Lopez-Santiago, R., Sanchez-Argaez, A. B., De Alba-Nunez, L. G., Baltierra-Uribe, S. L. & Moreno-Lafont, M. C. Immune response to mucosal Brucella infection. Front. Immunol. 10, 1759. https://doi.org/10.3389/fimmu.2019.01759 (2019).
-
Moreno-Lafont, M. C. et al. Antigen-specific activation and proliferation of CD4 + and CD8 + T lymphocytes from brucellosis patients. Trans. R Soc. Trop. Med. Hyg. 96, 340–347. https://doi.org/10.1016/s0035-9203(02)90119-7 (2002).
-
Clapp, B., Yang, X., Thornburg, T., Walters, N. & Pascual, D. W. Nasal vaccination stimulates CD8(+) T cells for potent protection against mucosal Brucella melitensis challenge. Immunol. Cell. Biol. 94, 496–508. https://doi.org/10.1038/icb.2016.5 (2016).
-
Schetters, S. T. T. et al. Outer membrane vesicles engineered to express membrane-bound antigen program dendritic cells for cross-presentation to CD8(+) T cells. Acta Biomater. 91, 248–257. https://doi.org/10.1016/j.actbio.2019.04.033 (2019).
-
Prior, J. et al. (ed, T.) Bacterial-Derived outer membrane vesicles are potent adjuvants that drive humoral and cellular immune responses. Pharmaceutics 13 https://doi.org/10.3390/pharmaceutics13020131 (2021).
-
Laidlaw, B. J. et al. Production of IL-10 by CD4(+) regulatory T cells during the resolution of infection promotes the maturation of memory CD8(+) T cells. Nat. Immunol. 16, 871–879. https://doi.org/10.1038/ni.3224 (2015).
-
Lee, J. B., Lee, K. A. & Chang, J. Phenotypic changes induced by IL-12 priming regulate effector and memory CD8 T cell differentiation. Int. Immunol. 19, 1039–1048. https://doi.org/10.1093/intimm/dxm072 (2007).
-
de Figueiredo, P., Ficht, T. A., Rice-Ficht, A., Rossetti, C. A. & Adams, L. G. Pathogenesis and immunobiology of brucellosis: review of Brucella–Host interactions. Am. J. Pathol. 185, 1505–1517. https://doi.org/10.1016/j.ajpath.2015.03.003 (2015).
-
Avila-Calderón, E. D. et al. Dendritic cells and Brucella spp. Interaction: the Sentinel host and the stealthy pathogen. Folia Microbiol. 65, 1–16. https://doi.org/10.1007/s12223-019-00691-6 (2020).
-
Labonte, A. C., Tosello-Trampont, A. C. & Hahn, Y. S. The role of macrophage polarization in infectious and inflammatory diseases. Mol. Cells. 37, 275–285. https://doi.org/10.14348/molcells.2014.2374 (2014).
-
Chen, S. et al. Macrophages in immunoregulation and therapeutics. Signal. Transduct. Target. https://doi.org/10.1038/s41392-023-01452-1 (2023).
-
Zuniga, E. I., Macal, M., Lewis, G. M. & Harker, J. A. Innate and adaptive immune regulation during chronic viral infections. Annual Rev. Virol. 2, 573–597. https://doi.org/10.1146/annurev-virology-100114-055226 (2015).
-
Tang, Y. et al. Serum levels of seven general cytokines in acute brucellosis before and after treatment. Infect. Drug Resist. 14, 5501–5510. https://doi.org/10.2147/idr.S341331 (2021).
-
Zhang, F. et al. The immunogenicity of OMP31 peptides and its protection against Brucella melitensis infection in mice. Sci. Rep. 9 https://doi.org/10.1038/s41598-019-40084-w (2019).
-
Fernandez-Prada, C. M. et al. Interactions between Brucella melitensis and human phagocytes: bacterial surface O-Polysaccharide inhibits phagocytosis, bacterial killing, and subsequent host cell apoptosis. Infect. And. immunity71, 2110–2119. https://doi.org/10.1128/IAI.71.4.2110-2119.2003 (2003).
-
Garcia-Quintanilla, M., Pulido, M. R., Pachon, J. & McConnell, M. J. Immunization with lipopolysaccharide-deficient whole cells provides protective immunity in an experimental mouse model of Acinetobacter baumannii infection. PLoS One. 9, e114410. https://doi.org/10.1371/journal.pone.0114410 (2014).
Acknowledgements
This study was supported scientifically by the Islamic Azad University Science and Research Branch.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Nobari, M.B., Razavi, M.R., Hosseini, F. et al. The Lpp-OmpA-BtaE fusion protein causes a protective immune response against Brucella melitensis in mice. Sci Rep 15, 27210 (2025). https://doi.org/10.1038/s41598-025-12059-7
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-12059-7