
Introduction
The extensive use of chemical herbicides in conventional agriculture has led to the evolution of herbicide-resistant weed species, particularly in response to the overuse of glyphosate. The repeated application imposes strong selection pressure, enabling weeds to develop and propagate resistance mechanisms to these substances1. One of the most problematic species is Chenopodium album, an annual weed widely distributed in North America that exhibits resistance to multiple herbicide classes, making it a persistent agricultural challenge in crop production systems2.
Beyond resistance, the environmental consequences of synthetic herbicide use are well-documented. Their continued application in common crops has been linked to soil and water contamination, adverse effects on non-target plants and animals, and the release of harmful compounds into the atmosphere during their production, transportation, and application3,4. These environmental drawbacks emphasize the urgency for transitioning to new weed management strategies that align with sustainability principles by developing herbicides derived from natural sources, commonly referred to as bioherbicides5. These agents, originating from bacteria, fungi, viruses, or plant extracts, have demonstrated considerable environmental benefits and are particularly suitable for integrated weed management6.
Plant-derived allelochemicals have attracted attention for their weed-suppressive potential and compatibility with regenerative agricultural systems. Several plant extracts have shown strong allelopathic effects, with recent studies highlighting their potential in integrated weed management7. Notably, methanolic extracts of Ocimum tenuiflorum and Prosopis juliflora, formulated into emulsifiable concentrates, have been shown to significantly reduce the density of Echinochloa colona, a dominant weed in rice cultivation, while also enhancing crop yield8. Within the Agavaceae family, Agave angustifolia and Agave cupreata have emerged as promising candidates for biocontrol applications, owing to their phytochemical-rich profiles that exhibit antifungal and possibly allelopathic effects9.
Based on these findings, Agave lechuguilla has emerged as a particularly relevant candidate for bioherbicidal research. Preliminary studies suggest its bioconverted extract inhibits seed germination, indicating an allelopathic potential10. As a native plant widely distributed across Mexico´s arid zones, A. lechuguilla is ecologically resilient and culturally and economically significant for rural and indigenous communities11. Traditionally harvested for “ixtle” (Tampico fiber), its fibrous residue, or bagasse, is typically discarded, representing an underutilized resource that could be revalued for agroecological innovation10.
Notably, while glyphosate is primarily characterized as a post-emergent herbicide, studies in glyphosate-resistant crops report reduced weed emergence when glyphosate is applied pre-sowing or pre-emergence, likely due to residual soil activity12,13. However, these effects are inconsistent across species and environments, and the molecular mechanisms, such as glyphosate’s interference with hormonal signaling or amino acid biosynthesis during germination, remain poorly understood. This gap highlights the need to compare glyphosate’s pre-emergent activity with novel bioherbicides, such as A. lechuguilla extract, which may offer more predictable, multi-target inhibition of weed establishment. Current literature offers adjacent insights that underline these deficiencies. For instance, recent studies have elucidated how seed-associated microbiomes promote germination through pathogen suppression and enzymatic degradation processes, suggesting that microbial and biochemical interactions may be crucial to understanding allelopathic effects at the molecular level14. However, such mechanistic depth remains unexplored for A. lechuguilla, particularly in comparison to well-studied herbicides like glyphosate. Moreover, research in allelopathy primarily emphasizes post-emergence herbicidal effects or the suppression of above-ground biomass. Conversely studies such as those on Trianthema portulacastrum highlight the emerging focus on pre-emergence impacts, particularly on seed germination inhibition. These gaps signal an urgent need for experimental studies that dissect the biochemical pathways of A. lechuguilla’s phytotoxicity and trials that isolate and test its pre-emergence activity against invasive or resilient weed species. Without these data, the development of A. lechuguilla-based bioherbicides will remain speculative, limiting their integration into agroecological practices tailored for arid and semi-arid environments.
In this study, we aim to revalorize the main residue from Agave lechuguilla as a potential pre-emergence bioherbicidal agent and fully harness its benefits over synthetic herbicides such as glyphosate. A key innovative aspect of our research is testing the hypothesis that the herbicidal activity of the natural extract operates through multiple modes of action. We validated this through transcriptomics and RT-qPCR, allowing for the comparison of differentially expressed genes, protein-protein interaction networks, and the identification of key hub genes. This investigation will enhance the understanding of AGE’s mechanisms and modes of action as pre-emergence herbicide, filling a gap in research that has predominantly concentrated on post-emergence action. Ultimately, this work can contribute to developing bioherbicides derived from plant extracts.
Results
The bioherbicidal activity of Agave lechuguilla extract compared with glyphosate
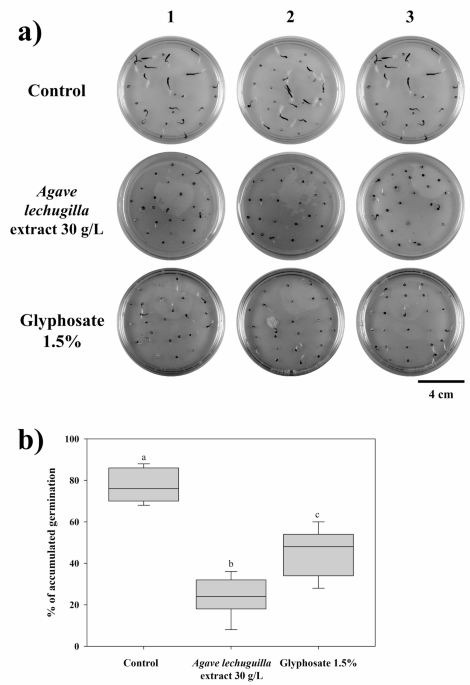
When the Agave lechuguilla extract was applied at a concentration of 30 g/L, it showed bioherbicidal activity against Chenopodium album seeds. Glyphosate was used as a control to assess its effects on inhibiting seed germination (Fig. 1A). The bioherbicide treatment significantly decreased germination compared to the control, and glyphosate was found to be less effective than the bioherbicide (Fig. 1B).

The bioherbicide activity on Chenopodium album seeds of Agave lechugilla extract. (A) Petri dishes containing MS media and seeds treated with 30 g/L of Agave lechugilla extract and 1.5% glyphosate, 1, 2, and 3 represent the experiment replicates. The Petri dishes illustrate the germination behavior, including differences in radicle emergence, hypocotyl elongation, and seedling orientation. (B) Boxplot showing the % of germination of each treatment (n = 25) after 72 h exposure. Different lower-case letters showed statistically significant differences determined through one-way ANOVA (p <.05), and the pairwise comparison was assessed with Duncan’s test (p <.05).
Transcriptome sequencing, assembly, and functional annotation of Chenopodium album
A total of 786,137 transcripts and 360,241 open reading frames (ORFs) were predicted from the transcriptome assembly. Both sets (nucleotide and protein sequences) were annotated using the BLASTx and BLASTp algorithms against the SwissProt database. The top three species distributions were correspond to Arabidopsis thaliana (31.4%), Homo sapiens (22.7%), and Mus musculus (17%) (Fig. 2A).

Functional annotation of Chenopodium album transcriptome. (A) Top species distribution of BLASTx and BLASTp against SwissProt database. (B) Gene ontology (GO) annotations, the % of transcripts were grouped in three different ontologies: biological process (BP) in blue, molecular function (MF) in red, and cellular component (CC) in green. (C) Clusters of Orthologous Genes (COG) annotations. Information storage and processing (yellow): [J] Translation, ribosomal structure, and biogenesis, [A] RNA processing and modification, [K] Transcription, [L] Replication, recombination, and repair [B] Chromatin structure and dynamics. Cellular processes and signaling (green): [D] Cell cycle control, cell division, chromosome partitioning, [Y] Nuclear structure, [V] Defense mechanisms [T] Signal transduction mechanisms, [M] Cell wall/membrane/envelope biogenesis, [N] Cell motility, [Z] Cytoskeleton, [W] Extracellular structures, [U] Intracellular trafficking, secretion, and vesicular transport, and [O] Posttranslational modification, protein turnover, chaperones. Metabolism (blue): [C] Energy production and conversion, [G] Carbohydrate transport and metabolism, [E] Amino acid transport and metabolism, [F] Nucleotide transport and metabolism, [H] Coenzyme transport and metabolism, [I] Lipid transport and metabolism, [P] Inorganic ion transport and metabolism, and [Q] Secondary metabolites biosynthesis, transport and catabolism. Poorly characterized (colorless): [R] General function prediction only, and [S] Function unknown. (D) KEGG Orthology (KO) annotations % of transcripts related to category metabolism (09100).
The gene ontology annotations comprised ~ 630,000 entries across three ontologies: biological processes (39.2%), molecular function (25%), and cellular component (35.8%). The top 10 annotations for each ontology are displayed in Fig. 2B. Among these, hormonal signaling terms (including auxin-activated signaling, cytokinin-activated pathway, and gibberellin biosynthesis) were enriched in the AGE-treated group, suggesting disruption of developmental regulation. KEGG analysis further revealed perturbations in flavonoid biosynthesis, glutathione metabolism, and MAPK signaling pathways, indicating oxidative and detoxification stress. Notably, the nitrogen compound metabolic process, biosynthetic process, and stress response each have at least 15,000 associated transcripts for biological processes. For molecular functions, ion binding, DNA binding, and RNA binding have over 10,000 transcripts. These biological processes and molecular functions are specifically located in organelles such as the nucleus, cytoplasm, and cytosol (see Fig. 2B).
For the Cluster Ortholog Genes (COG) annotations, a total of 3,012 annotations were obtained, divided into the following categories: information storage and processing (30.6%), cellular processes and signaling (18.4%), metabolism (51%), and poorly characterized (0%). Among these, the most abundant categories were translation, ribosomal structure, and biogenesis (19.5%), post-translational modification, protein turnover, and chaperones (5.7%), as well as amino acid metabolism (12.0%) (Fig. 2C).
A total of 49,616 transcripts were categorized in the KEGG Orthology annotation. This analysis focused specifically on metabolic pathways, with the following breakdown: carbohydrate metabolism accounted for 17.6%, amino acid metabolism for 17.5%, and lipid metabolism for 14.3% (Fig. 2D).

(A) Volcano plots illustrating differentially expressed genes are shown, with up-regulated genes in blue and down-regulated genes in red. The numbers represent differentially expressed genes with a log2 (fold-change) threshold > 2 and p-value < 0.001. (B) Venn diagram illustrating genes associated with abioteic stress in the presence of a bioherbicide.
Differentially expressed genes analysis in seeds exposed to Agave Lechugilla extract and glyphosate
Differentially expressed genes (transcript isoforms) were classified into three pairwise comparisons: (I) control vs. A. lechugilla extract, which identified a total of 3,246 genes; (II) control vs. glyphosate, totaling 5,234 genes; and (III) glyphosate vs. A. lechugilla extract, which included 6,240 genes (Fig. 3A). To compare the differentially expressed genes (DEGs) across the three groups, a Venn diagram illustrates the uniquely expressed DEGs as well as those shared among the groups. Group I has 539 unique DEGs and shares 292 with Group II and 2,269 with Group III. Group II contains 2,301 unique DEGs and shared 2,495 with Group III. Overall, the three groups share 146 DEGs (Fig. 3B). The magnitude of gene expression changes can be explored by examining the number of genes exhibiting different expression levels. These changes can be categorized into larger changes (> 5) and smaller changes (between > 2 and < 3) (Table 1). Suppl Table 1 shows Gene Ontology (GO) terms associated with the biological processes of the clusters depicted in the Venn diagram (Fig. 3A). This information helps us understand the GO terms related to the biological responses to both Agave lechuguilla extract and glyphosate.

The percentage of the pie charts indicates the number of GO terms associated with each cluster. (A) Up-regulated genes in 24 clusters. (B) Down-regulated genes into 5 clusters. (C) Up-regulated genes into 50 clusters. (D) Down-regulated genes into 15 clusters. (E) Up-regulated genes grouped into 18 clusters. (F) Down-regulated genes into 5 clusters.
Biological process GO terms clustering
Due to the redundancy of GO terms, clustering allows for a clearer overview of the most significant biological processes. In the comparison between control and Agave lechuguilla treatments, the clusters of up-regulated genes include negative regulation of biological processes (31.93%), response to oxygen-containing compound (12.61%), reproductive structure development (10.92%), biological regulation (10.92%), response to stimulus (5.88%), chemical response (5.04%), defense response (5.04%) and nitrogen compound metabolic processes (4.20%) (Fig. 4A). The down-regulated genes GO terms were classified into 5 clusters: Peroxidase activity (45.5%), photosynthesis, light harvesting in photosystem I (18.2%), actin polymerization or depolymerization (18.2%), and root hair elongation (18.2%) (Fig. 4B). In the comparison of control and glyphosate, up-regulated organonitrogen compound biosynthetic process (17.3%), cellular response to stress (12.7%) and regulation of RNA metabolic process (11%) can be seen (Fig. 4C), and down-regulated nucleic acid metabolic process (26.45%), plant-organ development (9.09%) and gene expression (7.44%) (Fig. 4D). The comparison between glyphosate and Agave lechuguilla extract for up-regulated is organonitrogen compound biosynthetic process (44.44%), response to inorganic substance (10.19%) and fruit development (8.33) (Fig. 4E) while down-regulated is gene expression (55.56%), reproductive system development (22.22%) and response to inorganic substance (11.11%) (Fig. 4F). All cluster information not shown can be found in Suppl Table 2.
Protein-protein interaction networks
To analyze differentially expressed genes, we constructed three protein-protein interaction networks: (i) control vs. Agave lechuguilla extract, (ii) control vs. glyphosate, and (iii) bioherbicide vs. glyphosate (Fig. 5). In group i), we identified 110 up-regulated genes, 15 down-regulated genes, and 39 genes predicted by GeneMANIA (Fig. 5A). In group ii), there were 109 up-regulated genes, 78 down-regulated genes, and 37 genes predicted by GeneMANIA (Fig. 5B). In group iii), we found 162 up-regulated genes, 37 down-regulated genes, and 29 genes predicted by GeneMANIA (Fig. 5C).

Predicted protein-protein interaction networks for differentially expressed genes. (A) Network 1 shows the up-and down-regulated genes in the control condition compared to the Agave lechuguilla extract condition. (B) Network 2 shows the up-and down-regulated genes in the control condition compared to the glyphosate condition. (C) Network 3 shows the up-and down-regulated genes between the Agave lechuguilla extract and glyphosate conditions. In all networks, blue triangles represent up-regulated genes, red inverted chevrons represent down-regulated genes, and gray circles indicate genes predicted by GeneMANIA. All networks are scale-free, and the interactions are based on biological process gene ontology (GO).
For Network 1 (Fig. 5A), the Gene Ontology (GO) terms for biological processes related to the down-regulated nodes indicate significant disruptions in various signaling pathways, including cell surface receptor signaling, gibberellic acid-mediated signaling, brassinosteroid-mediated signaling, and cytokinin-activated pathway. Developmental processes such as xylem development and embryo development, which lead to seed dormancy and flower development, are also affected. Additionally, metabolic processes are down-regulated, including biosynthetic pathways for L-histidine, inositol, L-phenylalanine, and tyrosine, as well as carbohydrate and cellulose catabolism and lipid metabolism (Suppl Table 2). The up-regulated nodes in Network 1 highlight enhanced biological processes. These include energy production, ATP synthesis driven by proton motive force, components of the electron transport chain, and the metabolism of 2-oxoglutarate, aspartate, and glutamate.
Additionally, the network indicates activation of various stress response pathways, including systemic acquired resistance mediated by salicylic acid, responses to insects, hypoxia, cold acclimation, jasmonic acid, and hyperosmotic conditions. In summary, the bioherbicide treatment triggered a complex response characterized by increased energy production and metabolism, activation of stress and defense pathways, alterations in development and reproduction, and changes in gene regulation and protein modification (Suppl Table 3). Notably, these interactions cluster around stress-induced hub genes rather than developmental regulators, differentiating AGE from glyphosate, primarily targeting transcription factors linked to meristem activity and organogenesis.
In Network 2, where the treatment is control versus glyphosate, the down-regulated genes indicate that glyphosate exposure deviates from complex responses involving detoxification, stress responses related to nutrient deprivation and oxidative stress, metabolic adjustments, and changes in gene expression, protein synthesis, and signaling pathways. In the case of up-regulated genes, the most prominent Gene Ontology (GO) terms are related to photosynthesis. These include photosystem II stabilization and assembly, phototropism, the photosynthetic electron transport chain, and light harvesting in photosynthesis. Significant GO terms associated with plant defense and stress response include systemic acquired resistance, salicylic acid-mediated signaling pathways, and response to herbivores (Suppl Table 4).
In Network 3, where the treatment involves a comparison between bioherbicide and glyphosate, the down-regulated genes associated with specific Gene Ontology (GO) terms indicate affected processes, including developmental processes, stem cell population maintenance, pattern specification, root meristem growth, root development, and seed dormancy. The signaling pathway impacted the ethylene-activated signaling pathway (see Supplementary Table 5). For the up-regulated genes, there are specific observations related to energy metabolism, cytoskeleton regulation, methylation, transport, stress response, and the development process (Suppl. Table 5).
Finally, we identified hub genes as those with the highest number of interactions in the networks, which are presented in Table 2. Some of these genes were selected to evaluate their relative expression by RT-qPCR (Table 3).
Relative mRNA expression analysis of Chenopodium album seedlings after Agave Lechuguilla extract and glyphosate exposure
Ten genes were selected to investigate the molecular mechanisms underlying the herbicidal effects of A. lechuguilla extract and glyphosate on Chenopodium album seeds, thereby validating the information obtained from the transcriptome differential gene expression analysis. Treatment with A. lechuguilla extract significantly up-regulated the aspartate peroxidase gene (CaAPX) in seeds compared to both control and glyphosate-treated seeds, with statistically significant differences (p <.01). The glyphosate treatment resulted in a significant down-regulation of the bifunctional purple acid phosphatase 26 gene (CabPAP2) and a significant up-regulation of the indole-3-acetic acid-amido synthetase gene (CaGH33) and the alpha carbonic anhydrase 8 gene (CaATCA8) in comparison to the control (p <.01). Notably, the expression of CaATCA8 was also significantly higher in Agave lechuguilla extract-treated plants than in those treated with glyphosate. These results suggest that glyphosate induces a different stress response in Chenopodium album seeds than the response elicited by Agave lechuguilla extract (Fig. 6A). The RT-qPCR results are presented as log2 (fold-change) to corroborate the differential gene expression patterns observed in the mRNA-seq experiment. The heatmap visualization highlights the up-and down-regulated genes in the 3 conditions (Fig. 6B). The RT-qPCR amplicons were analyzed on a 2% agarose gel to confirm the amplification of a single product and verify the expected molecular weight (Suppl Fig. 6).

Relative mRNA expression. The RT-qPCR results of ten selected differentially expressed genes. (A) The y-axis represents the relative mRNA expression (target gene/CaPPA2A). At the same time, the x-axis lists the ten selected genes: APX (ascorbate peroxidase), PAP26 (bifunctional purple acid 26), CAT (catalase), KSC19 (3-ketoacyl-CoA synthase 19), CBS (chlorophyll a-b binding protein CP26), ALS (acetolactate synthase, chloroplastic), GH33 (indole-3-acetic acid-amido synthetase), GLO5 (glycolate oxidase 5), APY (apyrase 2), and ATCA8 (alpha carbonic anhydrase 8). The bars indicate the mean, and the error bars represent the standard error (n = 9), the dashed line represents the normalized control treatment. An asterisk indicates statistically significant differences for each condition per gene, evaluated using the t-student test (p <.01). (B) Heatmap of relative mRNA expression in terms of log2 (fold-change).
Molecular mechanisms involved in bioherbicide activity of Agave Lechuguilla extract on Chenopodium album seeds
The molecular mechanisms that elucidate the bioherbicidal mode of action of Agave lechuguilla extract include the disruption of hormonal signaling, modulation of antioxidant enzymes, and metabolic suppression of phenylpropanoid and lignin synthesis. The specific processes influenced by AGE are illustrated in Fig. 7.

Summary of the molecular mechanisms of the bioherbicidal activity of Agave lechugilla extract on Chenopodium album seeds.
Discussion
This study aims to evaluate the molecular mechanisms underlying the bioherbicidal activity of an extract from Agave lechuguilla, which is derived from the bagasse (guishe) obtained during the extraction of ixtle (tampico fiber) from the community of Ramos Arizpe, Coahuila de Zaragoza, Mexico. A previous study demonstrated the effectiveness of bioherbicide of the extract, which was bioprocessed using fungal fermentation, on model seeds such as corn (Zea mays) and tomato (Solanum lycopersicum)10. This study evaluated the A. lechuguilla extract on the weed Chenopodium album without any prior bioconversion. This species is regarded as one of the five worst weeds in North America, and its management has largely relied on herbicides, resulting in a high level of resistance to these chemical treatments15,16. Previous research has remarked that non-chemical alternatives, such as using allelopathic compounds derived from agricultural residues, can effectively combat this weed16,17,18. While glyphosate is recognized for its post-emergence efficacy, it also impacts seedlings19. This study demonstrated that glyphosate has a pre-emergence effect on the seeds of C. album. It functions by inhibiting the enzyme 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS), which disrupts the synthesis pathway of the amino acids phenylalanine, tyrosine, and tryptophan20.
The bioherbicidal potential of A. lechuguilla extract against C. album seeds was demonstrated, revealing significant inhibition of germination in seeds treated with the extract. While glyphosate is predominantly utilized as a post-emergence herbicide, our findings indicate a comparable pre-emergence effect on C. album seed germination. These results underscore the significance of this study, as we seek to elucidate the specific molecular mechanisms involved in seed germination when exposed to both A. lechuguilla extract and glyphosate through transcriptomic analysis. While mRNA-seq provides a valuable overview of transcriptional responses and offers important insights into potential mechanisms of action, it is crucial to recognize that this technique has limitations. Specifically, it does not capture post-transcriptional regulation, protein modifications, or other layers of biological control. Nonetheless, studies like this are vital for generating hypotheses and directing future focused investigations aimed at fully understanding the mechanisms behind the bioherbicidal activity of Agave lechuguilla extract. This foundational work is a crucial step toward developing sustainable, plant-based herbicides for agricultural applications.
The raw data demonstrated high-quality sequencing, as we assembled over a billion paired-end reads. The quality of this assembly is confirmed by the mapping of 96% of the reads and further supported by the N50 and N10 values, which suggest a complete and representative assembly of C. album transcriptome. The obtained values provide a solid foundation for analyzing differential expression and functional annotation of the genes. The species distribution analysis revealed a high proportion of genes homologous to Arabidopsis thaliana, reflecting the close phylogenetic relationship between the two species. Gene Ontology (GO) annotation highlighted an overrepresentation of terms associated with metabolic processes, stress responses, and ion and DNA binding, particularly in the nucleus, cytoplasm, plasma membrane, and mitochondria. The Clusters of Orthologous Groups (COG) analysis identified the dominance of categories related to translation, amino acid metabolism, and energy production, indicating high metabolic activity.
KEGG Orthology (KO) annotations revealed that carbohydrate and amino acid metabolism are the most prominent metabolic pathways in gene expression. This result suggests a close relationship with the metabolic processes affected during C. album germination when exposed to A. lechuguilla extract and glyphosate. Overall, it is reported that exposure to xenobiotics like glyphosate can alter carbohydrate metabolism, which subsequently impacts amino acid metabolism21.
Differential expression analysis identifies differentially expressed genes (DEGs), classified as either down-regulated or up-regulated. The results revealed distinct DEGs in the control treatments, A. lechuguilla extract, and glyphosate in C. album. These treatments can affect various molecular functions and cellular processes. Additionally, some DEGs in the treatments may indicate both unique and shared modes of action (Fig. 3). The differentially expressed genes (DEGs) identified in the A. lechuguilla extract group, based on biological processes, indicate that the extract exerts an herbicidal effect by inducing cellular stress, altering carbohydrate metabolism, causing structural changes in the cell wall, and disrupting hormonal functions.
In contrast, the DEGs from the glyphosate group show changes in gene expression, protein modification, transport, metabolism, alterations in cell wall structure, hormonal signaling, and stress responses. Notably, both the A. lechuguilla extract and glyphosate share DEGs that involve alterations in cell wall structure, induction of oxidative stress, interference with hormonal signaling, and changes in gene expression (Suppl Table 1). These molecular changes are reflected in the germination results, where the seedlings that did germinate appear smaller in size and lack pigmentation (Fig. 1A). The herbicidal activity of A. lechuguilla extract can be attributed to the presence of various allelopathic compounds, which have been shown to induce bursts of reactive oxygen species in target plants, trigger programmed cell death, lead to lipid peroxidation, and cause changes in the cell membrane22,23. In the case of glyphosate, a thinning of the cell wall mediated by auxin accumulation has been reported in cotton (Gossypium hirsutum)24. Given these findings, it is essential to provide a thorough description of the GO avoiding redundant annotations, to gain a deeper understanding of the underlying molecular mechanisms.
To reduce redundancy, we conducted ClueGO analysis to generate clusters of specific terms, which helped simplify the results of the differential expression analysis. Our findings indicate that AGE leads to changes in nitrogen metabolism, activation of plant defense mechanisms, responses to chemical stimuli, and the development of reproductive structures. Conversely, we observed that the extract inhibits root growth, alters cell wall structure, and affects the photosynthetic process (see Fig. 4A and B). In the case of glyphosate, we found that it enhances nitrogen metabolism, cellular responses to stimuli, the regulation of RNA biosynthetic processes, and embryo development, ultimately contributing to seed dormancy. However, it also inhibits nucleic acid metabolism and organ development (see Fig. 4C and D). While there are differences between AGE and glyphosate, both treatments increase organonitrogen compound metabolic processes and responses to external stimuli, with a concurrent decrease in gene expression related to the development of reproductive structures and responses to chemical stimuli. Although both treatments appear to induce significant changes, some similarities in their modes of action regarding nitrogen metabolism are evident. It is important to note that glyphosate inhibits the EPSPS enzyme, crucial for synthesizing aromatic amino acids-precursors for various molecules and proteins. This inhibition disrupts nitrogen absorption and assimilation, leading to deregulation of carbon flow25,26. In the case of AGE, the effects on nitrogen metabolism appear quite broad. Oxidative stress can damage various structures, affect nutrient absorption and disrupt hormonal signaling. AGE is known to contain flavonoids, which can exhibit allelopathic activity. This activity may lead to alterations in germination, seedling growth, photosynthesis, and deregulation of the hormonal cascade and nutrient absorption23,27,28.
The protein-protein interaction networks provided a system-level perspective on how A. lechuguilla extract (AGE) and glyphosate disrupt cellular processes. The mapping of differentially expressed genes (DEGs) explores key hub genes and functional clusters, shedding light on the distinct mechanisms through which AGE and glyphosate affect C. album seeds. Network 1 (Fig. 5A) reveals a dual mechanism of herbicidal action characterized by hormonal and developmental disruptions and metabolic suppression. This mechanism is associated with down-regulated genes enriched in hormonal signaling pathways, including gibberellic acid, brassinosteroids, and cytokinin. These pathways involve processes such as xylem, embryo development, and germination.
On the other hand, metabolic suppression is associated with the biosynthesis pathways of specific amino acids (L-histidine and L-phenylalanine) and cell wall components, which impair structural integrity and nutrient assimilation (see Suppl Table 3). In response to these disruptions, a heightened response to stress includes energy production, cytoskeletal reorganization, and oxidative stress mitigation (see Suppl Table 3). The profile of AGE flavonoids, known for their allelopathic activity, includes several key compounds (flavanone, isorhamnetin, catechin, cyanidin, delphinidin, hesperidin, quercetin, apigenin, kaempferol, naringenin)23. While some allelopathic mechanisms of flavonoids, such as inducing oxidative stress, disrupting auxin transport, and inhibiting cell division, are well characterized27 many specific molecular targets remain unidentified. The bioactivity of AGE is likely due to its diverse flavonoid content, as previously suggested in profiling studies23. However, a definitive chemical fingerprint using analytical techniques like LC-MS or GC-MS is currently lacking. Future studies should correlate specific bioactive compounds with observed transcriptomic responses to enhance the precision and efficacy of bioherbicide formulation. Generally, it is known that these mechanisms may include inhibition of cell growth, disturbances in ATP production, hormonal disruption, and the generation of reactive oxygen species (ROS)22,29,30. Network 2 (Fig. 5B), we examine the genes involved in the control vs. glyphosate treatment. The analysis reveals that these genes are related to photosynthetic compensation and the disruption of detoxification pathways.
Additionally, we observed hormonal and metabolic suppression, followed by stress activation (Suppl Table 4). These processes align with the inhibition of the shikimate pathway and demonstrate effects on redox homeostasis and photoprotection25,31,32. Network 3 plays a crucial role in the analysis as it highlights the differences in the molecular responses of C. album seeds to aging (AGE) and glyphosate. This network emphasizes the distinct modes of herbicidal action associated with each condition. Seeds exposed to glyphosate experienced significant suppression of developmental processes, including stem cell maintenance, root meristem growth, ethylene signaling, and seed dormancy (Suppl Table 5). In contrast, aging (AGE) activated processes such as ATP metabolism, the electron transport chain, cytoskeletal organization, and stress acclimation (Suppl Table 5).
Network analysis has revealed that AGE can induce various forms of drought-related stress, as indicated by hyperosmotic stress, response to hypoxia, and jasmonic acid signaling (as seen in Network 1 and Network 2). Notably, the carbonic anhydrase gene (CaATCA8) showed a 30-fold increase in expression. This increase suggests an accumulation of atmospheric CO2 in seedlings, as it has been observed that this mechanism can alleviate drought stress by increasing pH levels through the presence of organic acids33. The chloroplastic acetolactate synthase (CaALS) gene also exhibited increased expression in the presence of AGE. This enzyme plays an important role in synthesizing branched-chain amino acids, which have been shown to help mitigate osmotic stress in plants34. Conversely, it has been noted that tolerance to drought stress is linked to stomatal regulation, with apyrase playing a crucial role. It has been observed that the gene encoding this enzyme shows increased expression in the presence of AGE and glyphosate35.
The increased expression of indole-3-acetic acid-amino acid synthetase (CaGH33) observed in both treatments suggests a common underlying mechanism with different hormonal disruptions. In the case of glyphosate treatment, it is known that the shikimate pathway is inhibited, which blocks the synthesis of tryptophan, a precursor to indole-3-acetic acid (IAA)36. This disruption could create a negative feedback mechanism for auxin regulation, as CaGH33 functions to detoxify excess auxins24. Conversely, lower expression levels of CaGH33 were observed in the case of AGE treatment; some of the flavonoid’s present could inhibit the transport of auxin inhibitors37,38.
Purple Acid Phosphatases (PAPs) are metalloenzymes that hydrolyze phosphate esters, releasing inorganic phosphate39. They may serve a dual role as both phosphatases and peroxidases and could be involved in regulating root development and senescence40,41. A decrease in PAP expression is observed when treated with glyphosate, while AGE is up-regulated. This activation might be related to the drought stress response, as suggested by other genes mentioned above42. On one hand, at alkaline pH, this enzyme can activate its peroxidase form and may be associated with the product of carbonic anhydrase, bicarbonate (HCO3−), which has an alkaline function39,43. On the other hand, this could serve as a counter-response to senescence, which can be induced by AGE in seeds41.
This study aims to identified the molecular mechanisms underlying the herbicidal effects of Agave lechuguilla extract compared with glyphosate on Chenopodium album seeds. The findings reveal both shared and unique pathways of action for each substance. The extract from Agave lechuguilla demonstrates bioherbicidal activity by inducing multi-target oxidative stress. The disruption of the redox balance affects energy metabolism, influences auxin signaling, and mimics drought-like responses in the plant. On the other hand, glyphosate kills plants primarily by inhibiting the enzyme EPSPS, which leads to metabolic paralysis, suppression of phosphate metabolism, and triggering compensatory photosynthesis, thereby disrupting auxin regulation and negatively impacting growth and development. This study highlights the potential of plant-derived herbicides (bioherbicides) as a solution for combating emerging chemical herbicide resistance while also supporting the goals of sustainable and regenerative agriculture. However, the findings are based on in vitro germination assays under controlled conditions. Field validation under realistic soil, climate, and microbial conditions is essential to translate these molecular insights into agronomic practice. Finally, these findings position Agave lechuguilla extract as a promising candidate for developing sustainable, plant-derived pre-emergent herbicides. Future studies focusing on formulation, field efficacy, and environmental impact assessment will be essential to advance this natural extract toward regenerative agricultural application. Besides, the focus of the research is to describe the basics of the herbicidal effect on a molecular level. The field validation is not in the core of the present state of the investigation; however, field validation is necessary. Future work should focus on formulating AGE for soil application and testing its efficacy in field trials against C. album and other resistant weeds.
Methods
Biological material and Agave lechuguilla extract
The Chenopodium album seeds were collected from San Rafael, Veracruz, Mexico, agricultural fields, although it is not an endangered or threatened plant, a collection permit was requested from the Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). The extract with bioherbicidal activity was prepared from the residue, known as “guishe” of the ixtle (Tampico fiber) industry from A. lechugilla in Cosme, Ramos Arizpe, Coahuila, Mexico. The collection of the waste was based on the official Mexican standard NOM-008-SEMARNAT-1996, which establishes that the waste from the use of Agave lechuguilla can be used without any authorization. The guishe was transported to the Environmental Biotechnology Laboratory of the Universidad Autonoma de Coahuila, where it was processed with a hydraulic press using 1.014 kN. The juice obtained was then processed in a household mixer model 62515RY (Proctor Silex) at maximum speed until it formed a dense foam. This material was placed in a tray dryer KL-10 (Koleff, Santiago de Queretaro, Mexico) at 50 °C. The resulting powder was stored in airtight plastic bottles for future use.
Chenopodium album collection and molecular identification
The Chenopodium album seeds were collected from weeds in agricultural in San Rafel, Veracruz, Mexico. The seeds were washed using 0.1% tween-20, followed by 3% sodium hypochlorite, and then rinsed with sterile distilled water five times. After soaking the seeds for 48 hours, they were placed in Petri dishes with Murashige-Skoog (MS) agar medium and allowed to germinate for 72 hours at a temperature of 25 °C with a photoperiod of 12 hours. The resulting seedlings were homogenated with the plastic pestle in a DNA extraction buffer44. The DNA was visualized in agarose gel with 0.8% electrophoresis and measured using spectrophotometric methods. The DNA isolated was utilized as a template on PCR to amplify the ITS region and a non-coding region of the chloroplast with the help of the following primers: forward 3’—CGAAATCGGTAGACGCTACG—5’ and reverse 3’—GGGGATAGAGGGACTTGAAC—5’45. The PCR reactions were performed using the GoTaq Flexi DNA Polymerase (Promega, Madison, WI) in a 25 µL reaction mixture containing: 1X Buffer Green GoTaq Flexi, 2.5 mM MgCl2, 0.2 mM dNTPs, 0.5 µM of each primer, and 1.25 U of Taq DNA polymerase. The temperature profile included an initial denaturation step at 95 °C for 2 min, followed by 35 cycles of denaturation at 95 °C for 30 s, annealing at 50 °C for 30 s, extension at 72 °C for 45 s, and a final extension step at 72 °C for 5 min. The PCR products were visualized using agarose gel electrophoresis and sequenced in both directions using Sanger sequencing. The obtained sequences were submitted to the NCBI and assigned the following accession numbers: PP668985, and PP658427. The seeds were deposited in the herbarium HCIB of the Centro de Investigaciones Biologicas del Noroeste, S. C. with the following accession number: HCIB32366. The molecular identification of Chenopodium album was conducted by A.E.V.-L. at CIBNOR facilities. Regarding Agave lechuguilla waste, it was identified by the members of the Ramos Arizpe Community as indicated by the NOM-008-SEMARNAT-1996.
Experimental design
Previous experiments assessed the bioherbicidal activity of A. lechugilla powder extract. The MS medium was prepared using 1.5% agar and sterilized. When the agar reached a temperature of 52 °C, the following enriched media were prepared: (1) MS medium only, (2) MS medium with A. lechuguilla powder (30 g/L) and (3) MS medium added with glyphosate (1.5%) for agricultural use in the form of isopropyl amine salt from the Arraza brand (Agrochemical Tridente, Coyoacán, Mexico City), gl. The media were poured into 100 × 15 mm Petri dishes. As detailed in the previous section, the C. album seeds were washed. They were then soaked for 48 h based on three different treatments: (1) control group with seeds soaked in distilled water only, (2) seeds soaked in A. lechugilla extract (30 g/L), and (3) seeds soaked in a 1.5% concentration of glyphosate, as recommended by the manufacturer for field use. The 1.5% glyphosate (isopropyl amine salt) concentration corresponds to standard agricultural use and for this experiment was used here as pre-emergent herbicide. Nonetheless, a direct molar equivalence to the AGE concentration (30 g/L) was not established, warranting further dose-response calibration to ensure fair efficacy comparisons. Following the soaking, 25 seeds were carefully placed in Petri dishes with their respective media and incubated at 25 °C for 72 h under a 12-hour light/dark photoperiod. Each treatment consisted of 9 replicates. The germination percentage was recorded and then calculated.
mRNA-sequencing of Chenopodium album seeds
The nine replicates per treatment were pooled into three biological replicates per treatment. The ungerminated seeds and seedlings were ground using liquid nitrogen and carefully transferred into 2 mL screw-cap tubes containing one ceramic zirconium-coated bead, 100 mg of glass bead (200–400 μm), and 600 µL RNA extraction buffer (100 mM Tris-HCl, 25 mM EDTA, 2.5% PVP, 2.5 M NaCl, 2.5% β-Mercaptoethanol). The powder-filled tubes were homogenized in FastPrep-24 at 4.5 m/s for 40 s. Subsequently, the tubes were cooled on ice, and the RNA extraction was performed following the protocol described by Vennapusa et al., 2020. The RNA was quantified using spectrophotometric methods and visualized in agarose (1.2%) gels to check the integrity of 18 s and 28 s rRNA bands. After that, the RNA samples were treated with DNAse I amplification grade (ThermoFisher Scientific, Waltham, MA, USA) as per the manufacturer’s instructions, and the mixture was purified using RNEasy (Qiagen, Germantown, MD, USA).
Azenta Life Sciences (South Plainfield, NJ, USA) performed the RNA library preparation and its sequencing. The RNA samples were quantified using a Qubit 2.0 fluorometer (ThermoFisher). The RNA integrity was assessed using 4200 TapeStation (Agilent Technologies, Palo Alto, CA, USA). The RNA sequencing library was prepared using the NEBNext Ultra II RNA Library Prep Kit for Illumina following the manufacturer’s instructions (New England Biolabs, Ipswich, MA, USA). Briefly, mRNA was initially enriched with Oligo(dT) beads, and the enriched mRNAs were fragmented for 15 min at 94 °C. First-strand and second-strand cDNA are subsequently synthesized. cDNA fragments are end-repaired and adenylated at 3’ends, and universal adapters are ligated to cDNA fragments, followed by index addition and library enrichment by PCR with limited cycles. The sequencing library was validated using the Agilent Tapestation 4200 (Agilent Technologies, Palo Alto, CA, USA) and quantified using Qubit 2.0 Fluorometer (ThermoFisher) as well as by quantitative PCR (KAPA Biosystems, Wilmington, MA, USA).
The sequencing libraries were multiplexed and clustered onto a flowcell. After clustering, the flow cell was loaded onto the Illumina HiSeq 3000 instrument according to the manufacturer’s instructions. The samples were sequenced using a 2 × 150 bp Paired-End (PE) configuration. The HiSeq Control Software conducted image analysis and base calling. Raw sequence data (bcl files) generated from Illumina was converted into fastq files and de-multiplexed using Illumina bcl2fastq 2.20 software. One mismatch was allowed for index sequence identification.
Transcriptome assembly, transcript quantification, and functional annotation
The mRNA-seq yielded 9 paired-end libraries, totaling 1,074,894,300 raw reads. These reads were de Novo assembled into 786,137 transcripts after being trimmed. The raw reads have been deposited in the Short Read Archive (SRA) database, and BioProject and BioSample have been created under accession number PRJNA1159996. Table 4 provides the key statistics derived from mRNA-seq and transcriptome assembly.
Each library was assessed using FastQC v0.12. Subsequently, the reads were trimmed using Trimmomatic v0.39 to remove adapters, discard low-quality reads below 3, and remove trailing low-quality reads below 3, and discard sequences shorter than 36 bases. Trimmed reads were assembled with Trinity v 2.15.1 using default parameters. The representation of the trimmed reads from each library in the transcriptome was then assessed using Bowtie2 v2.5.2. The quantification of each transcript abundance was assessed using the Trinity v2.15.1 pipeline; Salmon v1.10.2 was used to map reads versus transcriptome. The differential expression analysis was conducted with the DESeq2 package of Bioconductor v3.19, using 3 replicates for each treatment. Three sets of differentially expressed genes (DEGs) were identified: (i) control vs. Agave lechuguilla extract, (ii) control vs. glyphosate, and (iii) Agave lechuguilla extract vs. glyphosate. Functional annotation was performed using the Trinotate v4.0.2 pipeline, which involves BLAST with the UniProt database. The differential expression data and annotation data were built in the SQLite database. The ORFs were annotated on KAAS (KEGG Automatic Annotation Server) for KEGG annotations46. ClueGO v2.5.10 was used to predict biological process term clusters using the DEGs of every comparison for a non-redundant analysis for GO terms.
Protein-protein interactions prediction
The interaction prediction (protein-protein) is made by selecting up-and down-regulated genes for the treatments compared: (i) control vs. Agave lechuguilla extract, (ii) control vs. glyphosate, and (iii) Agave lechuguilla extract vs. glyphosate. The corresponding sequences (up- and regulated genes) were used to perform blast with the Arabidopsis thaliana database obtained from UniProt. The matching IDs were loaded into the GeneMANIA app in Cytoscape v3.10.2. The interaction networks were created to analyze up-and down-regulated genes based on various interaction levels, including gene co-expression, physical and genetic interactions, pathway participation, protein co-localization, protein domain similarity, and predicted protein interactions. The distribution of node degrees was used to identify the hub genes in each network, indicating the genes with the most interactions with their first neighbors. The hub genes were analyzed on the DAVID website.
Gene selection and primer design
Five reference genes were selected, the expression of which did not change in the three treatments evaluated, and ten genes of interest related to the molecular response of C. album seeds exposed to the Agave lechugilla extract. The primer sequences were designed in primer3plus with the next parameters: 60 °C Tm, 18–24 bp length, 70–250 bp amplicon size, 35–65%GC (Table 3).
Relative gene expression
To assess the efficiency of each primer, an aliquot of each cDNA sample was pooled and quantified in Nanodrop 1000 (ThermoFisher), and then serial dilutions were made at 1:10, 1:100, 1:1000, and 1:10,000. Carefully a master real-time PCR (qPCR) master mix was prepared with the following components: 1X colorless GoTaq Flexi buffer, 2.5 mM MgCl2, 0.2 mM dNTPs, 500 nM of each primer, 1X CES (0.54 M betaine, 1.34 mM DTT, 11 µg/mL BSA, 1.34% DMSO), 1X EvaGreen Dye (Biotium, Fremont, CA, USA) and 1.25 U GoTaq Flexi DNA polymerase (Promega, Madison, WI, USA). The qPCR reactions were carried out in the Rotor-Gene Q thermal cycler (Qiagen) in 0.1 mL strip tubes. The temperature profile used was 95 °C 5 min, a 45 times cycle of 95 °C 20 s, 60 °C 25 s and 72 °C 30 s, a melt curve of 55–99 °C (1 °C/s). The data were analyzed with Rotor-Gene Q software v2.1.0.9 (Qiagen).
The RT-qPCR reactions were performed under the conditions mentioned above with 3 technical replicates and 3 biological replicates for each treatment (same samples of the mRNA-seq). A 1:50 dilution of cDNA was used for gene amplification in each sample. To assess the stability of reference gene expression, five genes were evaluated: elongation factor 1-α (CaEFA1), glyceraldehyde-3-phosphate dehydrogenase (CaGAPDH), the catalytic subunit of serine/threonine-protein phosphatase PP2A-2 (CaPP2A2)47 ubiquitin-conjugating enzyme E2 W (CaUBE2), and the large ribosomal subunit protein uL2z (CaRPL8). Among these, CaPP2A2 exhibited the highest stability and was selected for calculating relative gene expression. Gene expression calculations were performed using the 2-ΔΔCq equation48. The statistical analysis was done using a t-student and assessing normality with the Kolmogorov-Smirnoff test (p <.05) in SigmaPlot v11 software (Systat Software Inc, Chicago, IL).
Compliance with guidelines
All experimental research involving Chenopodium album, including seed collection and molecular identification, complied with relevant institutional, national, and international guidelines. As this species is neither endangered nor protected, no formal ethical approval was required for its use in this study.
Data availability
The data pertinent to this research have been deposited in the NCBI repository ([PRJNA1159996](https://www.ncbi.nlm.nih.gov/bioproject/?term=PRJNA1159996)) and included in the supplementary materials. Additionally, the data supporting the findings of this study are available from the corresponding authors, [AGRA, and LJRG], upon reasonable request.
References
-
Riechers, D. E. et al. Herbicide resistance is complex: a global review of cross-resistance in weeds within herbicide groups. Weed Sci. 72, 465–486 (2024).
-
Bajwa, A. A., Zulfiqar, U., Sadia, S., Bhowmik, P. & Chauhan, B. S. A global perspective on the biology, impact and management of Chenopodium album and Chenopodium murale: two troublesome agricultural and environmental weeds. Environmental Science and Pollution Research 26, 5357–5371 (2019). (2019).
-
Mahmood, I., Imadi, S. R., Shazadi, K., Gul, A. & Hakeem, K. R. Effects of pesticides on environment. Plant, soil and microbes: 1. Implications Crop Sci. 253–269. https://doi.org/10.1007/978-3-319-27455-3_13/FIGURES/2 (2016).
-
Maria, M. G., da Silva, A. C., dos Santos Araújo, R. & Rigotto, R. M. Evaluation of the atmospheric contamination level for the use of herbicide glyphosate in the Northeast region of Brazil. Environ. Monit. Assess. 191, 1–11 (2019).
-
Pacanoski, Z. Bioherbicides. In Herbicides, Physiology of Action, and Safety Vol. 1 (eds Price, A. et al.) 253–274 (IntechOpen, Rijeka, 2015).
-
Hasan, M., Ahmad-Hamdani, M. S., Rosli, A. M. & Hamdan, H. Bioherbicides: an Eco-Friendly tool for sustainable weed management. Plants 2021. 10, Page 1212 (10), 1212 (2021).
-
Khamare, Y., Chen, J. & Marble, S. C. Allelopathy and its application as a weed management tool: A review. Front. Plant. Sci. 13, 1034649 (2022).
-
Salman, M. M. et al. Eco-friendly approaches for controlling Echinochloa colona in rice ecosystems using natural herbicides. PlScT 11, 5395 (2024).
-
Almazán-Morales, A. et al. Phytochemical profile and in vitro activity of Agave angustifolia and A. cupreata extracts against phytopathogenic fungi. Revista Mexicana De Fitopatología. 40, 169–187 (2022).
-
Sánchez Robles, J. H. et al. Initial Study of Fungal Bioconversion of guishe (Agave lechuguilla Residue) Juice for Bioherbicide Activity on Model Seeds. Fermentation 9, (2023).
-
Astier, M. et al. Back to the roots: Understanding current agroecological movement, science, and practice in Mexico. Agroecology Sustainable Food Syst. 41, 329–348 (2017).
-
Parker, R. G., York, A. C. & Jordan, D. L. Weed control in Glyphosate-Resistant corn as affected by preemergence herbicide and timing of postemergence herbicide application. Weed Technol. 20, 564–570 (2006).
-
Dogan, M. N., Ünay, A., Boz, Ö. & Ögüt, D. Effect of pre-sowing and pre-emergence glyphosate applications on weedsin stale seedbed cotton. Crop Prot. 28, 503–507 (2009).
-
Li, D. et al. Seed microbiomes promote Astragalus mongholicus seed germination through pathogen suppression and cellulose degradation. Microbiome 13, 23 (2025).
-
Van Wychen, L. & Weed Science Society of America National Weed Survey Dataset. Survey of the Most Common and Troublesome Weeds in Broadleaf Crops, Fruits & Vegetables in the United States and Canada. Weed Science Society of America National Weed Survey Dataset (2019). https://wssa.net/wp-content/uploads/2019-Weed-Survey_broadleaf-crops.xlsx
-
Eslami, S. V. & Ward, S. Chenopodium album and Chenopodium murale. Biology and Management of Problematic Crop Weed Species, 1st Edition 89–112 (2021). https://doi.org/10.1016/B978-0-12-822917-0.00009-4
-
Grimmer, O. P. & Masiunas, J. B. The weed control potential of oat cultivars. Horttechnology 15, 140–144 (2005).
-
Awan, I. U., Khan, M. A., Zareef, M. & Khan, E. A. Weed management in sunflower with allelopathic water extract and reduced doses of a herbicide. Pakistan J. Weed Sci. Res. 31, 19–30 (2009).
-
Schuster, C. L., Shoup, D. E. & Al-Khatib, K. Response of common lambsquarters (Chenopodium album) to glyphosate as affected by growth stage. Weed Sci. 55, 147–151 (2007).
-
Maeda, H. & Dudareva, N. The Shikimate pathway and aromatic amino acid biosynthesis in plants. Annu. Rev. Plant. Biol. 63, 73–105 (2012).
-
Serra, A. A. et al. Genome-wide transcriptional profiling and metabolic analysis uncover multiple molecular responses of the grass species lolium Perenne under low-intensity xenobiotic stress. Front. Plant. Sci. 6, 148892 (2015).
-
Šoln, K., Klemenčič, M. & Koce, J. D. Plant cell responses to allelopathy: from oxidative stress to programmed cell death. Protoplasma 259, 1111–1124 (2022).
-
Morreeuw, Z. P. et al. High throughput profiling of flavonoid abundance in Agave Lechuguilla Residue-Valorizing under explored Mexican plant. Plants 2021. 10 (Page 695 10), 695 (2021).
-
Yasuor, H. et al. Glyphosate-Induced anther indehiscence in cotton is partially temperature dependent and involves cytoskeleton and secondary wall modifications and auxin accumulation. Plant. Physiol. 141, 1306–1315 (2006).
-
Orcaray, L., Zulet, A., Zabalza, A. & Royuela, M. Impairment of carbon metabolism induced by the herbicide glyphosate. J. Plant. Physiol. 169, 27–33 (2012).
-
Duke, S. O. & Powles, S. B. Glyphosate: a once-in-a-century herbicide. Pest Manag Sci. 64, 319–325 (2008).
-
Weston, L. A., Mathesius, U. & Flavonoids Their Structure, Biosynthesis and Role in the Rhizosphere, Including Allelopathy. Journal of Chemical Ecology 39, 283–297 (2013). (2013).
-
Radhakrishnan, R., Alqarawi, A. A. & Abd Allah, E. F. Bioherbicides: current knowledge on weed control mechanism. Ecotoxicol. Environ. Saf. 158, 131–138 (2018).
-
Mierziak, J., Kostyn, K. & Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014. 19, Pages 16240–16265 (19), 16240–16265 (2014).
-
Parvez, M. M., Tomita-Yokotani, K., Fujii, Y., Konishi, T. & Iwashina, T. Effects of Quercetin and its seven derivatives on the growth of Arabidopsis Thaliana and Neurospora crassa. Biochem. Syst. Ecol. 32, 631–635 (2004).
-
de Freitas-Silva, L. et al. Glyphosate-induced oxidative stress in Arabidopsis Thaliana affecting peroxisomal metabolism and triggers activity in the oxidative phase of the Pentose phosphate pathway (OxPPP) involved in NADPH generation. J. Plant. Physiol. 218, 196–205 (2017).
-
Zobiole, L. H. S., Kremer, R. J., Oliveira, R. S. & Constantin, J. Glyphosate affects chlorophyll, nodulation and nutrient accumulation of second generation glyphosate-resistant soybean (Glycine max L). Pestic Biochem. Physiol. 99, 53–60 (2011).
-
Li, M. et al. Metabolomics analysis reveals that elevated atmospheric CO2 alleviates drought stress in cucumber seedling leaves. Anal. Biochem. 559, 71–85 (2018).
-
Huang, T. & Jander, G. Abscisic acid-regulated protein degradation causes osmotic stress-induced accumulation of branched-chain amino acids in. Arabidopsis Thaliana Planta. 246, 737–747 (2017).
-
Zhang, Y. et al. Populus euphratica apyrases increase drought tolerance by modulating stomatal aperture in Arabidopsis. Int. J. Mol. Sci. 22, 9892 (2021).
-
Lee, T. T. Mode of action of glyphosate in relation to metabolism of indole-3-acetic acid. Physiol. Plant. 54, 289–294 (1982).
-
Peer, W. A. & Murphy, A. S. Flavonoids and auxin transport: modulators or regulators? Trends Plant. Sci. 12, 556–563 (2007).
-
Du, H. et al. A GH3 family member, OsGH3-2, modulates auxin and abscisic acid levels and differentially affects drought and cold tolerance in rice. J. Exp. Bot. 63, 6467–6480 (2012).
-
Schenk, G., Mitić, N. Š., Hanson, G. R. & Comba, P. Purple acid phosphatase: A journey into the function and mechanism of a colorful enzyme. Coord. Chem. Rev. 257, 473–482 (2013).
-
Bhadouria, J. & Giri, J. Purple acid phosphatases: roles in phosphate utilization and new emerging functions. Plant Cell Reports 2021 41:1 41, 33–51 (2021).
-
O’Gallagher, B. et al. Arabidopsis PAP17 is a dual-localized purple acid phosphatase up-regulated during phosphate deprivation, senescence, and oxidative stress. J. Exp. Bot. 73, 382–399 (2022).
-
Wilkinson, S. PH as a stress signal. Plant. Growth Regul. 29, 87–99 (1999).
-
Lewandowska, J. & Laurent, J. P. Changes in apoplastic ph and membrane potential in leaves in relation to stomatal responses to CO2, malate, abscisic acid or interruption of water supply. Planta 213, 594–601 (2001).
-
Zhang, Y. P., Uyemoto, J. K. & Kirkpatrick, B. C. A small-scale procedure for extracting nucleic acids from Woody plants infected with various phytopathogens for PCR assay. J. Virol. Methods. 71, 45–50 (1998).
-
Taberlet, P., Gielly, L., Pautou, G. & Bouvet, J. Universal primers for amplification of three non-coding regions of Chloroplast DNA. Plant. Mol. Biol. 17, 1105–1109 (1991).
-
Kanehisa, M., Furumichi, M., Sato, Y., Matsuura, Y. & Ishiguro-Watanabe, M. KEGG: biological systems database as a model of the real world. Nucleic Acids Res. 53, D672–D677 (2025).
-
Xie, F., Wang, J. & Zhang, B. RefFinder: a web-based tool for comprehensively analyzing and identifying reference genes. Funct. Integr. Genomics. 23, 1–5 (2023).
-
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta delta C(T)) method. Methods 25, 402–408 (2001).
Acknowledgements
We would like to express our gratitude for the grant “Pp F003 11/II-O/2023” from the National Strategic Programs (ProNaCes) of Secretaria de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI), which was awarded to LJRG and AGRA, as well as the postdoctoral fellowship (Estancias Postdoctorales por México) #4493288 to AEVL. We would also like to thank the members of the Environmental Biotechnology Laboratory at the Universidad Autonoma de Coahuila, especially Gustavo Alberto Neyra Escobedo, M. Sc., for his technical assistance. Additionally, our thanks go to the laboratory technicians of CIBNOR, Delia Irene Rojas Posadas, M. Sc., María Sofía Ramos Galvan, Dr. Alfonso Medel Narvaez and Dr. Julio Antonio Hernández González. To Dr. Gracia Alicia Gómez Anduro for the support for RT-qPCR equipment. Lastly, we extend our thanks to Gilberto Méndez Griffin, who collected the seeds of the Chenopodium album in the agricultural field.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Velázquez-Lizárraga, A.E., Rios-González, L.J., Montiel, L.G.H. et al. Transcriptomic profiling reveals Agave Lechuguilla extract as a multi-target pre-emergent bioherbicide candidate against Chenopodium album. Sci Rep 15, 35796 (2025). https://doi.org/10.1038/s41598-025-17383-6
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-17383-6