
- Article
- Open access
- Published:
- Karpaga Raja Sundari Balachandran1 na1,
- Greeshma Mani1 na1,
- Akhil Thayyil Sidharthan2,
- J. T. Mary Leema1,
- Roobamathi Senthilkumar1 &
- …
- Dharani Gopal1
Scientific Reports volume 15, Article number: 35782 (2025) Cite this article
Subjects
Abstract
Deep-sea encompasses a wide diversity of microbiomes including bacteria, fungi and viruses which play crucial significant roles in nutrient biogeochemical cycling thereby imparting majorly to functional biodiversity of these hotspots. Sea mounts harboring microbes with extremophilic properties found in deep oceans could be conserved as living repository by functional metagenomics approach which is a potent source to screen bioactive compounds and novel enzymes thereby could address biological question on developing next generation plant biostimulants. This study outlines construction of fosmid metagenome library and adapted combined strategy of functional and nanopore sequence-based metagenomic screening to unveil phosphatase enzymes from Arabian Sea seamount sediment. About 9068 metagenomic clones were generated with an average insert size of 38 kb and stored in pools of 1024 clones, out of which 42 were found to be positive for phosphatase. Five clones with high phosphatase activity were further characterized and NIOT F41 showed the greatest specific activity for phosphatase (41.2 U/mg). Gluconic (1041 mg/L), oxalic (327 mg/L), and succinic acids (610 mg/L) were the predominant organic acids produced by recombinant clones. Fosmid DNA were extracted from five potential clones for nanopore-based metagenomics sequencing which generated an average of 6,00,786 reads. Taxonomic analysis revealed an abundance of Proteobacteria and Firmicutes phyla harboring phosphate-solubilising bacteria Pseudomonas aeruginosa, Bacillus subtilis and Staphylococcus warneri. Furthermore, functional annotation using phosphorus cycling database (PCycDB) predicted variation in relative abundance of phosphatase gene clusters encoding alkaline phosphatase (PhoD, PhoX and PhoA) and acid phosphatase (OlpA, PhoNand PhoC) produced by recombinant clones. In the pot assay, potential metagenomic clones exhibited positive impacts on shoot length (9.1 ± 1.1 cm, p < 0.05), root length (2.05 ± 0.05 cm, p < 0.05), wet biomass (39.3 ± 0.65 mg, p < 0.05), and dry biomass (5.1 ± 1.15 mg, p < 0.05) compared to the negative control indicating significant effect on promoting plant growth. The advanced nanopore sequencing and functional metagenomics methods employed in this study could serve as a marine biodiversity conservation approach for deep-sea microbes hidden in sea mount sediments towards harnessing potential next generation plant biostimulants with promising biotechnological application for sustainable agriculture.
Similar content being viewed by others
Introduction
The field of metagenomics has been revolutionized by technological advancements in synthetic biology, bioinformatics, and sequencing which could help to unravel microbiomes and their functional interaction in deep-sea ecosystem with significant bio-prospection perspectives. Functional and sequence-based metagenomics have been recently adopted towards understanding microbiomes of different ecological niche and bio-prospecting novel biocatalysts from uncultured microbes in environmental sources including water and soil1. Recent studies proposed that sequence-based metagenomics could enable the identification of potential genes involved in biogeochemical cycling, whereas function-based screening involves the development of clones from metagenomic libraries of various environmental niche to bio-prospect enzymes with a specific function2. Hence, it would be essential to combine both approaches for conservation of deep-sea microbiome through developing functional metagenome libraries of extreme environments, which would result in preserving biodiversity hotspots and better characterization of their enzymes for industrial application.
Oceans, as a habitat, provide a complex and extraordinary physical and chemical environment that is essential for all types of microorganisms, particularly marine bacteria, viruses and protists to thrive in specific ecological niche3. Deep-sea sediments, within marine habitats, constitute a harsh environment for microbial populations due to high pressure, low temperature, lack of light, poor nutrient availability and are recognized as well adapted versatile environment with understudied oceanic ecosystems, which imparts the significant necessity for conservation strategies in this fragile ecosystem to ascertain their future sustainability4. Seamounts are typically regarded as hotspots of species diversity and oases of biomass abundance due to their distinct ecosystems in the deep ocean5,6. Considerable efforts have been undertaken in the last few years to investigate the diversity, ecology, and function of the prokaryotes that live in seamount habitats through the use of metagenomics7, which has made it possible to record the diversity of microbes and investigate their functional potential in the ocean, where more than 99% of microbes remain yet uncultured8. Recent bathymetric surveys report on many of the selected seamount region in the Laxmi basin of Arabian Sea indicating their volcanic origin, which suggests that sea mounts of Arabian Sea could be potential hotspots for bio-prospection of industrially important enzymes from marine microbes with extremophilic properties9,10. Hence, it is proposed that metagenomic clone libraries could be used for conserving unculturable microorganisms as living repository of Indian deep-sea seamounts samples for a longer period towards addressing solutions through exploring sustainable biotechnological applications from recombinant microbes without exploiting the Oceanic environment for routine sampling and collection of samples.
A microbial community as a whole can be treated as a super- or mega-organism, whose metabolic pathways and networks are accessible with the use of metagenomics approaches11. Metagenome sequencing analysis has emerged as a potent technique for comprehending the biogeochemical cycling (e.g., phosphorus, nitrogen, carbon, sulphur, and metals) driven by microbes in different environments3,12. Oxford Nanopore Technologies is a powerful metagenomics tool, producing long reads that resolve complex genomic regions like repetitive or conserved elements which effectively recovers complete metagenome assembled genomes from various samples, including sludge, water, sediments, and feces, demonstrating its versatility13. Its ability to deliver fast results at a lower cost makes it an invaluable tool for diverse applications, from agricultural studies to medical research14. Nanopore sequencing based method has recently emerged as significant metagenomics technique, which has been implemented in our earlier studies towards understanding microbial diversity and harnessing potential of deep-sea environment including Bay of Bengal15 and Arabian Sea sediment16. Several detailed studies have been conducted on the cycling and fractionation of phosphate in the deep ocean of Arabian Sea. Recent research showed that the formation of a coastal upwelling zone in the Southeastern Arabian Sea during southwest and northeast monsoons results in the cycling of Phosphorus17. The genes responsible for phosphorus cycling (PCGs) and the microorganisms involved in the process are not thoroughly studied18. Alkaline phosphatases (EC 3.1.3.1) are the most investigated phosphoric monoester hydrolases and its genes phoD and phoX are shown to be more prevalent in marine environments than phoA19,20,21. The gene encoding a new alkaline phosphatase was discovered in a metagenomic library constructed from ocean-tidal flat sediments of Korea’s west coast which was identified to be a part of a newly discovered PhoX family, different from the well-researched classical PhoA family22. There has been limited research on the gene clusters that play a role in the cycling of phosphorus in deep-sea sediments, leaving a significant gap in our understanding of this crucial process.
Recently, several sequencing techniques have been applied in soil metagenomes to predict the entire sequence of biosynthetic gene clusters in silico23,24. Metagenomic methods have also been adopted to clone and sequence soil genome for screening specific enzymes7 where its functional analysis could lead to more accurate metagenome annotation than sequence-based analysis alone25. Functional screening of enzymes through metagenomics could find potential implication to address the challenges associated with low enzyme expression and complex purification processes26. The most appropriate vector must be chosen to insert the metagenomic DNA depending on the objective of functional screening wherein expression vectors like fosmids, cosmids (< 40 kb), bacterial artificial chromosomes (BACs), and yeast artificial chromosomes (YACs) (> 40 kb) are useful for larger biosynthetic gene clusters27,28. Recent reports unveil that soil metagenomes could be cloned using cosmid or fosmid library to identify the biosynthetic gene clusters efficiently29 which is adopted in the present study for constructing metagenomic libraries from Arabian Sea seamount sediments.
In general, it is necessary to confirm functional screening of any enzyme from metagenome clone library by sequencing-based methods30,31. Recent reports on sequencing of fosmid clone libraries with large inserts revealed that functional screening of fosmids clones with specific enzyme activity must be de novo sequenced which is more significant towards identifying specific gene targets31. Earlier reports reveal that numerous novel genes that encode variant enzymes from marine environments have been discovered by functional metagenomics32, which include lipase from marine sponge33, cellulase from Coral, Siderastrea stellata34 and DNA polymerase from glacial ice35. Fosmid metagenome library from ocean tidal flat sediments has been previously reported to have undergone fluorescence-based screening for lipase, cellulase, and phosphatase through GESS-FACS22,36. Recent progress in functional metagenomics screening methods combined with advances in next generation sequencing technologies could result in efficient bio-prospection of significant enzymes and metabolites from specific environmental niche31. These coupled methodologies have been adopted for mining industrial biocatalyst esterase from deep-sea samples including thermal water of alkaline hot spring at Lobios (Ourense, Spain)37 and Atlantis IIbrinepool, Red Sea(Indic Ocean)38 even amylase from Arabian Sea sediment39,40.
Several recent reports reveal the significance of Phosphate-solubilizing bacteria (PSB) as resourceful biofertilizers for sustainable agriculture due to its ability to secrete alkaline phosphatase (phoD)41, acid phosphatase (phoC)42 and phytase enzymes43 that can solubilise insoluble phosphorus in the soil for efficient absorption, uptake and utilization by plants44. Currently, only a small number of such plant growth-promoting bacteria are used in agriculture45,46 wherein our recent findings revealed two potential phosphate solubilising bacteria Priestia megaterium and Bacillus velezensis, from seamount sediments of Arabian Sea which promotes plant growth47. Recent approaches target the use of synthetic synergistic communities, obtained from one or multiple soil microbiomes that show best growth conditions even under biotic or abiotic stress, aiming at a broader target range, higher robustness and a better plant-beneficial efficacy48,49. Furthermore, advanced nanopore sequencing methods is emerging as a potent tool to generate plant biostimulants that could help to increase crop yield50, thereby resulting in development of next generation fertilizers to overcome harmful impact of synthetic molecules on environment51. However, there are no reports on bio-prospection of plant biostimulating enzymes from biodiversity hotspots including deep-sea sediment which could serve as a living metagenomic repository of extremophilic enzymes and biomolecules that could be used for sustainable agriculture with implication in environmental restoration.
This study aimed to bio-prospect phosphatase enzyme using functional metagenomics approach by constructing a metagenomics clone library using fosmid vector from seamount sediments of Arabian Sea as methodology to develop biological innovations through conservation of deep-sea microbes. Further, the potential metagenomic clones producing maximum phosphatase were sequenced by nanopore long read technology to understand the microbiome hidden in metagenomic clones of dark seamount sediment and to confirm functional potential of PSB towards identifying phosphate cycling gene expression variation conferring phosphate solubilisation. This study focused to correlate the abundance of PSB from seamount sediment with plant growth promoting properties towards understanding its implication in sustainable agriculture.
Materials and methods
Collection of sample
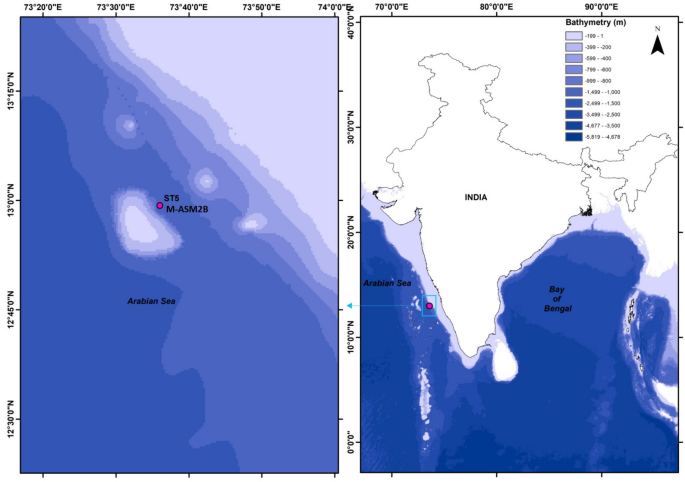
Marine sediment samples were collected from Arabian Sea on board NIOT-Ocean Research vessel, Sagar Manjusha (SAMA 10) cruise in November 2022. About 30 g of sediment was collected and transferred to a labelled sterile bag, then stored at 4 °C on board before being transferred to − 40 °C at the laboratory facility for further analysis. This study focused on Arabian Sea sediment collected from bottom of seamount M-ASM2B (Station 5–12.988333°N, 73.600283°E) at a depth of 850 m. Bathymetric mapping for the sample location of station 5 (ST5) was illustrated using ArcGIS 10.8 and bathymetric data acquired from https://gebco.net/ (Fig. 1). Environmental DNA from this sediment sample was utilized to construct a fosmid metagenomic library, subsequently screened and sequenced to characterize the phosphatase enzyme from the recombinant clones.

Bathymetric mapping of seamount showing sample location (ST5- M-ASM2B) collected at a depth of 850 m in Arabian Sea. The map was generated using ArcGIS Desktop (ArcMap) version 10.8 (esri, https://www.esri.com/en-us/arcgis/products/arcgis-desktop/overview) and the sampling location is indicated by a red dot which is the bottom of a seamount.
Construction of fosmid metagenomic library from seamount sediment
High molecular weight environmental DNA extraction method was adopted from Patin and Goodwin52 and further modified for seamount sediment samples to yield high quality DNA, where optimized lysis buffer was used for extraction53 followed by Polyethylene glycol (PEG-6000) for precipitation of high molecular weight DNA54. Multiple extractions were carried out and pooled together to attain a higher concentration of metagenomic DNA for cloning. The quality of DNA was analysed using agarose gel electrophoresis and quantified using Nanodrop Spectrophotometer (DS-11 Spectrophotometer, Denovix). The extracted DNA was purified using a silica-based column as per the kit protocol (Qiagen DNeasyPowerSoil Pro kit). Metagenomic library was constructed using Copy Control Fosmid Library Production Kit with pCC1FOS Vector from Epicentre Biotechnologies (CopyControlFosmid Library Production Kit, Lucigen, USA). Five micrograms of extracted metagenomic DNA were end-repaired and size selected according to the manufacturer protocol. The required amount of DNA was ligated, packed in lambda phages and were transformed into EPI300 Escherichia coli cells55. The recombinant clones were selected on LB agar plates with chloramphenicol (12.5 µg/mL). The clones obtained were stored as pooled clones in 20% glycerol at -80 °C in 96 well plates for further functional screening and sequencing of potential clones.
Confirmation of recombinant clones
To confirm the insertion of metagenomic DNA in the recombinant clones, fosmid DNA was extracted from the random clones by alkaline lysis method with some modifications56. Fosmid DNAs were purified with 2.5 M Ammonium acetate precipitation followed by ethanol wash and eluted in nuclease-free water. Restriction digestion was carried out with Not I enzyme (10 U) in fosmid DNA (500 ng) to confirm the size of inserted metagenomic DNA in the clones. PCR amplification of fosmid DNA was also carried out using pCC1FOS Sequencing Primers (as suggested by the manufacturer), 5′-GGATGTGCTGCAAGGCGATTAAGTTGG-3′ (Forward) and 5′-CTCGTATGTTGTGTGGAATTGTGAGC-3′ (Reverse) to confirm the insert size of metagenomic DNA. PCR amplification was carried out using fosmid DNA (200 ng), PrimeSTAR GXL DNA Polymerase (1.25 U) from Takara which is specifically used to amplify long amplicons. PCR reaction cycle was carried out in 50 µL reaction volume on 96 well thermocycler: initial denaturation was carried out at 95 ⁰C for 5 min, 35 cycles of amplification as follows: denaturation at 95 ⁰C for 30 s, annealing at 50 ⁰C for 30 s, elongation at 72 °C for 30 s and final elongation was performed at 72 °C for 5 min. The size of the PCR products was analyzed using agarose gel electrophoresis.
Screening of phosphatase enzyme from metagenome clone library
The fosmid metagenome library could be screened for phosphatase using either function-based screening or sequence-based screening. Function-based screening helped to identify the potential metagenomic clones producing required enzymes using a specific substrate, while sequence-based screening locates the biosynthetic gene clusters in the library and efficiently identify the microbiome present in metagenomic DNA. The methodology for combination of functional and sequence-based screening approach followed in the present study for bio-prospecting phosphatase enzyme from metagenome clone libraries of Arabian Sea sediment is illustrated in Supplementary Fig. 1.
Agar based primary screening for phosphatase
The pooled clones were subjected to substrate-based screening for phosphatase enzyme on NBRIP agar media containing tricalcium phosphate (TCP) as substrate57 and bromophenol blue as pH indicator which was incubated at 37 °C for 48 h. The recombinant clones with halozones around it were selected as phosphate solubilisers with phosphatase activity.
Quantification of phosphatase
Phosphatase enzyme was quantified for the positive clones using p-nitrophenyl phosphate (p-NPP) as substrate by spectrometric method. The recombinant clones with halozones were inoculated in 200 µL of NBRIP broth in 96 well plates while EPI300 E. coli served as negative control. The plate was incubated at 37 °C for 48 h. The phosphatase activity of recombinant clones was measured with a colorless substrate p-nitrophenyl phosphate (p-NPP), which yielded yellow p-nitrophenol product upon phosphate solubilisation (p-NP)58. The reaction mixture consisted of 200 µL 1 mM p-NPP (dissolved in 0.1 M Glycine-NaOH buffer) and 200 µL of cell free extract, made up to a fixed final volume of 1300 µL with nuclease free water. After a reaction time of 60 min at 25 °C in dark, the reaction was stopped by adding 200 µL of 0.05 M NaOH. The generated product p-nitrophenol (p-NP) was measured by spectrophotometry (UH5300 Spectrophotometer, HITACHI) at 405 nm59. The concentration of phosphate released was estimated from the standard graph of para-nitrophenol (p-NP). GraphPad Prism 7 and SPSS 29.0 were used, respectively, for the graphical depiction and statistical analysis of phosphatase activity of recombinant clones. Test data were subjected to a one-way ANOVA using SPSS 29.0 whereas differences between means at p < 0.05 were evaluated by Tukey HSD test.
Extraction of phosphatase enzyme and its partial purification
The potential recombinant clone with maximum phosphatase activity was selected for extraction of phosphatase enzyme. The selected clones were inoculated in specific media containing peptone 5 g; yeast extract 2.5 g; glucose 1 g; Mg2SO4 0.05 g; K₂HPO₄ 0.2 g; distilled water 1 L60 and incubated at 37℃ for 2 days to estimate the protein content. The cells were suspended in 50 mM Tris HCl buffer (pH 7.4), sonicated for 10s, and then cooled on ice for 15 s. This cycle was repeated 10 times after which the cells were collected at 10,000 g for 15 min. The cell free supernatant was precipitated by 80% saturated ammonium sulfate and dialyzed (membrane cut off 12,000Da, Sigma) against the same buffer at 4 °C overnight61. Using bovine serum albumin (BSA) as a standard (Sigma, Germany), the protein content of crude and partially purified phosphatase was quantified62. The activity of phosphatase enzyme in both the crude and partially purified protein was assayed using p-nitrophenyl phosphate (p-NPP) and measured spectrophotometrically (UH5300 Spectrophotometer, HITACHI) at 405 nm59.
Estimation of organic acid
Organic acid quantification was performed for the five potential recombinant clones with maximum phosphatase activity using a Shimadzu HPLC system with a quaternary pump, auto-sampler, and programmable UV-Vis detector. Organic acid separation was performed using a Phenomenex Luna c-18 column (4.6 *150 mm, 5 μm particle size), protected by a guard column (Phenomenex, c-18, 5 μm particle size) and detected at 214 nm. The data was collected in three dimensions – absorbance, time, and wavelength – using LC Solutions software. A 100% isocratic mobile phase of 50 mM potassium phosphate buffer with a pH of 2.8063 was used. The flow rate was set to 0.5 mL/min, and the column was kept at 35 °C using a column oven. Prior to injection, the sample and standard were passed through a 0.22 μm PTFE syringe filter (Acrodisc, Pal German, Germany). Calibration curves were developed for organic acid standards, which include lactic acid, oxalic acid, succinic acid, citric acid, gluconic acid, acetic acid, and formic acid.The maximum phosphate solubilising clones were inoculated in 30 mL NBRIP broth medium. The flasks were incubated at 37 °C in an orbital shaker at 100 rpm for 48 h. About 5mL of bacterial culture was centrifuged at 10,000 rpm for 15 min and filtered through 0.22 μm nylon membranes (acrodisc, Pal German, Germany) to obtain cell-free supernatant. Twenty microlitres of filtered supernatant were injected into the HPLC system (LC2010, Shimadzu). Identification and quantification of organic acids in the recombinant clones were performed by comparing retention times and peak areas of respective standards (Supplementary Table 1). All the samples were analyzed in duplicates.
Sequencing of potential recombinant clones encoding maximum phosphatase
Fosmid DNA was extracted from the five phosphatase positive clones using same methodology as described. The concentration and purity of DNA was measured using Nanodrop Spectrophotometer (DS-11 Spectrophotometer, Denovix). The integrity of DNA was observed on agarose gel electrophoresis and the DNA concentration was quantified using Qubit dsDNA HS assay kit in Qubit fluorometer (Qubit 4 Flourometer, Invitrogen Thermo Fischer Scientific). DNA samples were end-repaired (NEBnext ultra II end repair kit, New England Biolabs, MA, USA), cleaned up with 1X AmPure beads (Beckmann Coulter, USA). Native barcode ligation was performed with NEB blunt/ TA ligase (New England Biolabs, MA, USA) using SQKNBD114.96 and cleaned with 1xAmPure beads. Qubit quantified barcode ligated DNA samples were adapter ligated for 15 min using NEBNext Quick Ligation Module (New England Biolabs, MA, USA). Library was cleaned up using 0.6X AmPure beads (Beckmann Coulter, USA) and finally sequencing library was eluted in 15 µL of elution buffer and sequenced on Nanopore PromethION system, (PromethION P24 and Data Acquisition Unit, ONT, Oxford, UK) using PromethION flow cell (FLO-PRO114M) according to effective library concentration and data amount required. NGS library preparation and sequencing was carried out at Genotypic Technology Pvt. Ltd., Bangalore, India.
Sequence pre-processing and taxonomic analysis
Nanopore sequenced reads were de-multiplexed using Guppy v2.3.4 and the quality was verified using Nanostat tool64. Data pre-processing includes removal of adapters, de-replication of reads and quality control65. Metagenome Assembly was performed using the Flye tool with meta-option and the processed high-quality reads were assembled into contigs. Contigs were compared against NT Database (non-redundant nucleotide seq) using Centrifuge36, which contain reference sequences from a wide range of organisms, including bacteria, Archaea, viruses and eukaryotes classifying from kingdom to higher taxonomic ranks. Pavian tool was further used to estimate and interactively explore the taxonomic content66. Sankey plot is used for representing taxa abundance and connections between different taxonomic ranks. Comparative taxonomic analysis was carried out using relative abundance values for top 10 taxa at species level which are represented by heat map generated using R-statistical vegan package Non-negative Matrix Factorization (NMF)67.
Functional annotation for phosphate cycling genes
Functional annotation was carried out using KEGG database68, where three level classifications was carried out to predict the number of reads involved in the enzyme hydrolysis. Further pathway annotation was carried out using PCycDB which is recently developed database specifically for prediction and fast analysis of phosphorus cycling genes from metagenome datasets18. The genes involved in oxidative phosphorylation, transporters, two-component system, and organic phosphoester hydrolysis are major processes for microbes to regulate, transport, and uptake P sources from the environment which were predicted from PCycDB using BLAST analysis18. R-statistical package NMF was used to generate heat map based on gene expression count for each samples.
Bio-priming and pot assay for plant growth application
Bio-primed tomato seeds (5 seeds per pot, in duplicates) were sown in each experimental pot of sterilised garden soil. This pot experiment was conducted over a 14-day period in a plant house located near the Department of Marine Biotechnology at the National Institute of Ocean Technology in Chennai where temperature was maintained between 25 ºC and 36 ºC which is optimal for plant growth. Throughout the study, humidity levels were kept between 75% and 80%, and standard daylight duration was measured at 12 h, with a light intensity ranging from 600 to 1000 µmol/m2/s. Fosmid recombinant cultures were sprayed to their respective pots, while double distilled water served as the negative control, and Paenibacillus pseudetheri (a phosphobacterium obtained from Manidharma Biotech Private Limited in Chennai) was used as the positive control. After 14 days of growth, the plants were uprooted to measure vegetative parameters such as root length, shoot length, and the weight of both roots and shoots (measured for both fresh and dry weight).
Results
Fosmid metagenomic library construction from seamount sediment sample
Metagenomic DNA of high concentration was obtained from seamount sediment of Arabian Sea using modified metagenomic DNA extraction method and visualised by agarose gel electrophoresis, which showed an intense band approximately 40 kb as shown in Fig. 2a. The quantification results indicated high molecular weight DNA of concentration1026 ng/µL with low A260/A280 value (1.4) and A260/A230 value (0.9) suggesting that some contaminants still remained in the modified protocol. The extracted DNA was purified using QIAGEN Power Soil Pro Kit (Qiagen Inc., USA) wherein purified DNA was recovered with concentration of 903.3 ng/µL, A260/A280 value of 1.8 and an A260/A230 value of 1.9 indicating its suitability for constructing fosmid metagenomic libraries. About 571 ng of end-repaired metagenomic DNA was used for ligation and its subsequent transformation in host cell E.coli which resulted in 9068 clones with an efficiency of 1.5 × 104 clones per µg of DNA (Fig. 2b). The library was constructed with an average insert size of 40 kb comprising about 362 Mbp of metagenomic DNA from the Arabian Sea seamount sample. These recombinant clones were stored in 20% glycerol as 1024 pooled clones which would enable efficient functional screening of enzymes from large number of soil metagenomics clones29.

(a) Metagenomic DNA Isolated from deep Arabian Sea sediment sample: 1—1 kb plus ladder, 2—λ DNA (48 kb), 3—eDNA extracted from M ASM2B (b). Recombinant clones of M-ASM2B selected on Luria Bertani Agar plate with chloramphenicol.
Extraction of fosmid DNA and confirmation of recombinant clones
Fosmid DNA from randomly selected metagenomic clones was extracted to confirm the efficiency of cloning. Alkaline lysis method followed by purification yielded a fosmid DNA of intact quality. Cloning efficiency was confirmed by restriction digestion and amplification of fosmid DNA. Four hours of restriction digestion of fosmid DNA with restriction enzyme NotI released 3 fragments as confirmed through gel electrophoresis wherein one of band was found to be approximately 40 kb in size, indicating the presence of metagenomic DNA in the recombinant clones. Fosmid DNA was extracted from random clones and amplified using PCC1FOS sequencing primers and long Taq polymerase which resulted in high molecular weight amplified product of approximately 40 kb indicating successful insertion of metagenomic DNA into the vector and demonstrating cloning efficiency of fosmid vectors(Supplementary Fig. 2).
Functional screening and quantification of phosphatase in recombinant clones
Out of 1024 pooled clones, 42 clones showed halozone formation around colonies on NBRIP with bromophenol blue agar plates indicating its phosphatase activity by primary screening method (Supplementary Fig. 3). All those 42 clones were considered positive for phosphatase production and further quantified for phosphatase enzyme by spectro-photometric method. Compared to EPI300 E. coli (negative control), all recombinant clones showed significantly increased phosphatase activity. Among them, NIOT F41 (330.06 µM/mL, p < 0.001) exhibited maximum phosphatase activity followed by NIOT F42 (277.93 µM/mL, p < 0.001), NIOT F10 (272.89 µM/mL, p < 0.001), NIOT F13 (261.11 µM/mL, p < 0.001) and NIOT F24 (230.85 µM/mL, p < 0.001) (Fig. 3). These five potential recombinant clones with maximum activity were selected for further downstream analysis.

Quantification of phosphatase using p-nitrophenyl phosphate (p-NPP) as a substrate measured in 42 recombinant clones. Legends show different color for various recombinant clones. The results shown are the mean values of replicates and the vertical bars represent standard errors (S.E.). All the data shown are statistically significant with p < 0.001.
Extraction and partial purification of phosphatase
The enzyme concentration was measured using para-nitrophenyl phosphate (pNPP) as substrate for both crude and partially purified enzyme and the total protein content was also measured to determine the specific activity of the phosphatase enzyme. In the present study, it is evident that partially purified phosphatase enzyme from NIOT F41 exhibited the maximum specific activity of 41.2 U/mg (Table 1). On the other hand, the specific activity of the crude enzyme for the same clone NIOT F41 was lower (5.2 U/mg) compared to partially purified enzyme. It was also observed that fold of purity was increased for partially purified enzyme of NIOT F41 (7.95) compared to crude enzyme (1.0). These results indicate that specific activity of the enzyme increased after partial purification by dialysis method in all the five potential recombinant clones.
Estimation of organic acid produced by recombinant clones
The organic acids produced by the recombinant clones were identified by HPLC from the standard curves of pure organic acids (Supplementary Table 1). Among these, three different organic acids were detected in higher concentration (gluconic acid, oxalic acid and succinic acid) by the recombinant clones as shown in Fig. 4. Gluconic acid was produced in higher concentration by NIOT F41 (637 mg/L), NIOT F13 (1041 mg/L) and NIOT F24 (997 mg/L). Oxalic acid and succinic acid were produced in higher concentration by the recombinant clones NIOT F42 (327 mg/L) and NIOT F10 (610 mg/L) respectively. A very few unknown acids were also detected, but none of them did match with the standards used for the identification. These results suggest that the production of organic acid could be attributed to the phosphate solubilisation by these recombinant clones.

HPLC chromatogram of recombinant clone (a) NIOT F10 showing peak for Succinic acid and Gluconic acid (b) NIOT F13 showing peak for Gluconic acid (c) NIOT F24 showing peak for Gluconic acid (d) NIOT F 41 showing peak for Gluconic acid (e) NIOT F42 showing peak for oxalic acid and Gluconic acid.
Pre-processing and assembly of sequenced fosmid reads
Fosmid sequencing was carried out for the five potential positive clones which showed synthesis of maximum phosphatase enzyme in the quantification analysis. Sequencing of five clones resulted in an average of 6,00,786 reads using long read sequencing technology and further processed to remove adapter sequence which generated the high-quality processed data for microbiome identification. The cloning vector sequence of pCC1FOS (Genbank accession EU140751) matching with the processed reads and also with the E. coli genome (NC_000913) were trimmed and removed by Deconseq tool (version 0.4.3) [filter parameters- 90% coverage and 94% identity]65. The assembled reads had an average contig length of 25,104 bp at an average of 1259 contigs across the sample and detailed statistics is represented in Table 2. The processed reads were used in further downstream analysis for taxonomic and functional annotation of all the samples.
Sequence annotation for taxonomic analysis
Taxonomic analysis of the processed reads was performed against centrifuge indexed NT database. The assignment of individual reads to taxa based on their hits were analyzed and summarized from kingdom to species level along with read counts (Supplementary Fig. 4). Taxonomic analysis of the five phosphatase positive clones revealed that the major phylum was found to be Firmicutes for NIOT F10 (50.82%) and NIOT F42 (81.82%), while Proteobacteria was represented higher in NIOT F41 (72%), NIOT F13 (71.28%) and NIOT F24 (84.72%). The bacterial classification from kingdom to species level is shown as Sankey vvisualization plot for each sample which revealed that class Bacilli was found abundant in NIOT F10 (50.88%) and NIOT F42 (81.86%) at class level (Supplementary Fig. 4a). On the other hand, class Gammaproteobacteria was represented high in NIOT F24 (80%), NIOT F41 (66.85%) and NIOT F13 (39.63%) as shown in Supplementary Fig. 4b. Among the genus, Pseudomonas was found to be abundant in NIOT F41 clone (64.86%) and NIOT F24 (78.7%), while Bacillus was found to be relatively higher in NIOT F10 (30.12%) and Staphylococcus was comparatively abundant in NIOT F42. At the species level, Pseudomonas aeruginosa was found to be majorly represented in three of the sequenced clones, NIOT F24 (43.46%), NIOT F41 (37.74%) and NIOT F13 (25.16%) whereas Bacillus subtilis (17.77%) and Staphylococcus warneri (43.46%) was highly represented in NIOT F10 and NIOT F42 respectively. Taxonomic analysis of microbiome in sediment metagenome results suggest that major genus including Pseudomonas, Bacillus and Staphylococcus identified from Nanopore sequencing of potential phosphatase producing clones harbored already reported phosphate solubilising bacterium in literature59,69.
Comparative taxonomic analysis
The relative abundance of taxonomic classification by heat map results confirmed that different phylum was majorly represented among the five clones indicating distinct properties of each clone with insertion of DNA from various organisms in the metagenomic DNA. The results of heat map generated at species level (Fig. 5) clearly showed that Paenibacillus sp. Y412MC10 and Bacillus subtilis was found to be relatively high in NIOT F10, while Pseudomonas aeruginosa was abundant in NIOT F13, NIOT F24 and NIOTF41 which were previously reported to be phosphate solubilising microbes in earlier studies59,69. On the other hand, Staphylococcus warneri was found to be represented high in NIOTF42 suggesting that all these five potential fosmid clones harbor phosphate solubilising microbes attributing to their maximum phosphatase activity59,70. Statistical diversity analysis results showed that among the five clones, Shannon index for NIOT F10 was found to be high (2.79) indicating diversity richness in this specific clone compared to others. On the other hand, Simpson index representing the diversity measure within a single ecosystem or sample was calculated as value close to 1 for all the five samples (Supplementary Table 2) indicating that there are several species in the community, and also the proportion of species population is evenly composed of all the representative species.

Heat map analysis showing species abundance in the five metagenomic clones. Different colour variation represents the relative abundance of the species across all five recombinant clones.
Functional classification
Major gene ontology classifications in all samples were represented in gene ontology distribution at level three (Supplementary Fig. 5). KEGG pathway analysis results showed that metabolism category was highly represented in all the five clones indicating their role in phosphate metabolism (Excel S1). Three level classifications were carried out by KEGG analysis for enzymes involved in phosphate cycling pathways. Hydrolases enzyme class was analyzed in detail since the phosphatase enzyme belongs to this class of enzymes (Excel S2). PCycDB analysis was carried out for functional analysis of phosphate cycling genes responsible for cellular P metabolic processes. Comparative analysis of gene expression count for phosphate cycling genes across five samples showed that each clone was unique wherein phoD and phoX gene that encode for alkaline phophatase was high in NIOT F24 while PhoA was relatively high in NIOT F13 (Fig. 6). On the other hand, different cluster of genes involved in synthesis of non-specific acid phosphatase, olpA and phoN was relatively abundant in NIOT F41 and NIOT F10 respectively but the major cluster of gene phoC encoding for same enzyme was produced high in NIOT F42. Notably, opd gene which encode for phosphotriesterase was majorly represented with high intensity in NIOT F13. Similarly, phy encoding for Phytase was also high in NIOT F13 but appA encoding for same enzyme was found to be very high in NIOTF10. These results suggest that the gene for alkaline phosphatase was encoded high in recombinant metagenome clones of NIOT F13 and NIOT F24 whereas the gene cluster for acid phosphatase were abundant in NIOT F41, NIOT F10 and NIOT F42 metagenomic clones. On the contrary, the genes encoding for phytases are high in NIOT-F13 and NIOT-F10 clones suggesting the unique characteristics of each clone for enhanced phosphatase activity (Fig. 6).

Heat map showing the gene expression count among five potent recombinant clones of Arabian Sea conferring genes related to phosphate cycling predicted by PCycDB analysis. Different genes encoding same class of enzymes are grouped and represented in similar color; yellow for alkaline phosphatase, green for acid phosphatase, blue for phosphor-triesterase and magenta for phytase.
Seed priming and pot assay for Plant growth application
Seed priming assay using tomato seeds treated with fosmid recombinant clones NIOT F10, NIOT F13, NIOT F24, NIOT F41 and NIOT F42 showed 70%, 70%, 60%, 80% and 80% seed germination, respectively, which were greater than that of negative control (50%) which is represented in Table 3. Whilst seeds primed with positive control showed 70% which is lower than two clones (NIOT F41 and NIOT F42) indicating the efficiency of metagenomic clones from Arabian Sea. In the pot assay, NIOT F24 exhibited statistically significant effect on shoot length, measuring 9.1 ± 1.1 cm (p < 0.05), which is 1.44 times longer than that of negative control. In contrast, the highest root length was observed in NIOT F10 (2.05 ± 0.05 cm, p < 0.05), representing a 1.46-fold increase compared to the negative control. Furthermore, plants treated with the recombinant clones showed a significant increase (p < 0.05) in both wet and dry biomass relative to the negative control. Notably, the wet biomass was highest in NIOT F10, at 39.3 ± 0.65 mg, which corresponds to a 1.99-fold increase compared to negative control. Moreover, the dry biomass of NIOT F42, reaching 5.1 ± 1.15 mg, was significantly higher than that of the negative control, showing an increase of 1.52-fold (Table 3). These results indicate that all the recombinant clones were shown increased plant growth attributes than negative control and comparable results with positive control.
Discussion
Metagenome libraries aid in the recovery and expression of biosynthetic gene clusters71. It is reported that metagenome libraries with large insert size might contain polycistronic genes where the complete metabolic pathways could be analyzed and studied72. Even though several vectors are available for cloning of large insert, fosmid vectors are the most prominent one which can produce a large number of clones by inserting poly-cistronic genes thereby increase the screening of functional molecules73. Fosmid vector is already reported as an effective vector for the metagenome library construction from ocean tidal flat sediment and water column22,74. As the metagenome fosmid library discussed in this study was constructed from an extreme environment, it was anticipated that novel biocatalysts with diverse biochemical and physiological characteristics would be bio-prospected from Arabian Sea seamount libraries.
The construction of large insert environmental metagenome libraries relies on the effective recovery and purification of high-quality environmental DNA. Although many extraction procedures have been developed to isolate nucleic acids directly from environmental samples, the post-extraction purification procedures can often be time-consuming and laborious75. In this study, modified metagenomic DNA extraction followed by silica-based column purification yielded high quality DNA. A metagenome fosmid library consisting of 9068 recombinant clones was constructed from environmental DNA of Arabian Sea seamount sediment. Reports highlight the need to conserve fragile ocean ecosystems4 emphasising the need for protecting biodiversity in extreme environment, which was carried out in the present study to unveil the importance of preserving the genetic diversity of microbial communities of ocean seamount sediments. Hence, fosmid metagenomic library constructed from Arabian Sea seamount could serve as a genetic resource or living repository enabling the utilization of functional genes of extremophiles even if the original microbial population becomes threatened or inaccessible in the future.
A recent study identified marine microbial genomes as a key resource for gene mining offering significant potential for discovering novel enzymes76, which was earlier reported as significant method for unraveling biocatalysts including lipolytic enzymes, proteases, oxido-reductases, glycosyl hydrolases, phosphatases/phytases from unexplored environment of soil and water1. Following prior findings on enzyme screening from fosmid clones, this study envisaged to screen phosphatase enzyme from metagenomics clone libraries of seamount sediment. One effective way to identify desirable enzymes from metagenomic libraries that are expressible in the cloning host is by activity-based plate screening method77 which was implemented in the present study for functional screening of metagenomics libraries. Prior studies on biosynthetic gene cluster identification from soil metagenome were carried out in pools of 2000 clones from a total of 83,700 recombinant fosmid clones indicating the efficient functional screening using pooled clone strategy29, which was adapted in this study to effectively screen a large number of clones because it is challenging to screen every recombinant clone for specific enzymes with desired activity. In this study, about 9068 clones were stored in pools of 1024 clones as glycerol stocks for efficient functional screening of enzymes and biomolecules from seamount habitats. Primary agar-based functional screening resulted in the identification of 42 recombinant clones from a total of 1024 pooled clones exhibiting phosphatase activity by halozone formation around colonies indicating their ability to solubilise phosphate from the media. Earlier studies reported about positive fosmid clones for phosphatase through agar-based screening from functional metagenome libraries of decomposing leaf litter78 and also in hydrocarbon-polluted soil79. Furthermore, alkaline phosphatase synthesised by phosphate solubilising bacteria has also been reported in our recent study from deep-sea sediment samples of Arabian Sea47. These studies support the rationale behind prospection of phosphatase enzyme from the metagenomic library of Arabian Sea seamounts.
The positive clones underwent further quantitative screening for phosphatase activity using pNPP as a substrate. pNPP has already been reported as a screening substrate for phosphatase in the fosmid library constructed from the metagenome of tidal flat sediments22. In this study, the maximum activity was shown by the metagenomic clone NIOT F41 (330.06 µM/mL, p < 0.001) which is higher than the activity shown by already reported PSB Pseudomonas aeruginosa from Nigeria80. These results suggest the potential of recombinant clones from Arabian Sea sediment for enhanced phosphatase activity which could be useful in phosphate solubilisation for agricultural soil81.
In this study, partially purified phosphatase enzyme from NIOT F41 recombinant clone showed the highest specific activity of 41.2 U/mg. Comparing this finding to similar studies from Serratia sp. exhibited a specific activity of 9.58 U/mg82 and Metarhizium anisopliae with 8.45 U/mg83, both of which was found to have lower specific activity than that observed in the recombinant clones from seamount sediment. The maximum specific activity exhibited by the clone NIOT F41 might be due to the combination of extremophilic PSBs in the fosmid clones of Arabian Sea sediment. Notably, it was observed that specific activity of clone NIOT F41 was 5.2 U/mg for crude enzyme when compared to 41.2 U/mg for the partially purified enzyme. The present findings were found to be consistent with the previous reports where the purified enzyme was found to demonstrate better specific activity than the unpurified one84. According to Razali et al.85, increasing enzyme purity leads to an increase in specific activity which suggests that antagonist enzymes such as proteases are eliminated during purification allowing the protein of interest to be in the best possible form to produce its actual catalytic activity.
According to Prasad et al.86, the primary mechanism forsolubilization of inorganic phosphates is through the action of organic acids. The release of soluble orthophosphate from tricalcium phosphate (TCP) by microorganisms typically involves the production of organic acids87,88. In order to solubilize inorganic phosphate compounds, PSBs have been found to produce some organic acids such as acetic, formic, lactic, gluconic, glycolic, oxalic, succinic, malic and maleic acid89. In the present findings, gluconic acid was found in higher concentration for NIOT F41 (637 mg/L), NIOT F13 (1041 mg/L) and NIOT F24 (997 mg/L). It was observed that oxalic acid and succinic acid were produced in higher concentration by the recombinant clones NIOT F42 (327 mg/L) and NIOT F10 (610 mg/L) respectively. According to the findings of Behera et al.90, PSB isolated from mangrove soil produced highest concentration of oxalic acid (289 mg/L), followed by succinic acid (0.5 mg/L), acetic acid (0.4 mg/L), malic acid (0.3 mg/L), and citric acid (0.2 mg/L). Organic acids production by PSB strains isolated from Moroccan mines was investigated by Mardad et al.91in NBRIP broth culture, where results revealed that gluconic acid was the majorly produced organic acid by all PSB strains, reaching concentrations of up to 83.287mM. These results of previous reports on organic acid production by PSBs are in congruence with the present findings on recombinant clones producing gluconic acid as the major organic acid during phosphate solubilization. These organic acids are reported to aid in phosphate solubilization thereby enhance plant growth in vitro92,93.
Fosmid sequencing using Nanopore technology is reported to be a cost-effective, high-throughput approach that facilitates the rapid discovery of enzymes31. Function based screening for phosphatase was carried out to identify clones showing maximum phosphatase activity which was confirmed by sequencing of five potential recombinant clones in Nanopore platform towards prediction of phosphate cycling genes encoded by the metagenomic clones. Nanopore-based metagenome sequencing has proven effective in recent studies towards assessing microbial community composition in agricultural soil which helped to correlate specific taxa with its functional properties94. Our study applied this long read sequencing method to explore the bacterial taxa involved in phosphate solubilisation from Arabian Sea towards understanding its plant growth promoting properties. Taxonomic findings from metagenome sequencing of five positive fosmid clones revealed major representation from Pseudomonas, Bacillus and Staphylococcus genera. These results were in congruence with previous studies which revealed that most dominant phosphate solubilizing bacterial genera were Pseudomonas, Bacillus, Vibrio, Alcaligenes, Micrococcus, Corynebacterium and Flavobacterium isolated from deep-sea sediments in the Bay of Bengal which contributed significantly to biogeochemical cycles and benthic productivity in the Exclusive Economic Zone (EEZ)95. Earlier studies have also shown that Staphylococcus was also found to be best phosphate solubilizing bacteria in addition to Bacillus, Pseudomonas and Paenibacillus isolated from tropical rainforest soil59,69. These results attribute to the fact that bacterial genus identified from sequencing of seamount sediment metagenomic clones provides strong evidence with earlier studies on PSB from different ecological nichewhich would be useful to develop plant biostimulants for sustainable agriculture.
Further insight into the species level classification revealed key phosphate-solubilising bacteria, including Pseudomonas aeruginosa, Bacillus subtilis, Paenibacillus sp. Y412MC10, and Staphylococcus warneri. Among this, P. aeruginosa has been identified as a phosphate solubilizer from Ganga River96, while B. subtilis and P. aeruginosa have been recognized as phosphate-solubilizing rhizobacteria97 and also exhibit stress-tolerant activity in agricultural fields98. Further, S. warneri was also found to solubilize tricalcium phosphate in oil palm rhizospheres in Colombia70. These results implicate that the species identified from metagenomic clone libraries of Arabian Sea sediment also involve in phosphorus solubilization and could enhance plant growth in phosphate deficit soil.
Recent reports reveal that two-component system, oxidative phosphorylation, transporters, and organic phosphoester hydrolysis are major processes for microbes to regulate, transport, and uptake P sources from the environment whereinphosphoester hydrolysis pathway was emphasized in this study to understand various cluster of genes encoding acid and alkaline phosphatase enzymes18. Among the phosphate cycling genes, phoA, phoD, and phoX encode for alkaline phosphatases while phoN, aphA, phoC, and olpA encode acid phosphatases. On the other hand, opd gene encodes phosphor-triesterase, while phy and appA code for phytases as reported in PcycDB, a database recently reported for prediction of these genes from metagenomic datasets18. Comparative taxonomic analysis of five potential metagenomic clones revealed that Pseudomonas aeruginosa was found to be majorly represented in NIOT F24 and functional analysis of phosphate cycling genes by heat map results showed that phoD, and phoX encoding for alkaline phosphates were relatively high in same clone. Literature studies of several Pseudomonads also showed conservation of key pho regulators in Pseudomonas aeruginosa, Pseudomonas putida and Pseudomonas aureofaciens where PhoD and phoX was found to be expressed under Pho regulon control99. On the other hand, Bacillus was found to be relatively abundant in NIOT F13 where comparative analysis also demonstrated that PhoA gene was found to be abundant in same clone, which could be correlated with reports on Bacillus subtilis for synthesizing alkaline phosphatase (PhoA) and various other enzyme and act as cell factory for agriculture, medicine and biomaterials100. A recent study emphasizes the complex interactions between microbial species in sediment communities and their cooperative role in nutrient cycling within deep-sea ecosystems101. This is relevant to the current study utilizing fosmid clones derived from microbial communities of the Arabian Sea seamount for understanding the phosphate cycling genes. Thus, the functional genes identified from microbiome of the seamount can be used for understanding biogeochemical cycles of marine hotspots.
Recent studies on tomato seeds treated with plant growth promoting bacterial strains revealed significant increase in germination102 which was correlated to current findings of seed priming results where highest germination index was shown by tomato seeds treated with recombinant clones NIOT F41 and NIOT F42 compared to negative control. Pot Assay results for Plant growth application showed that shoot and root length of tomato seedlings were also highest in the recombinant clones than positive control and other plant growth attributes showed comparable results with positive control. There are earlier studies on significant plant growth when tomato seeds treated with tri-species consortia of PSBs including B. gladioli, Pseudomonas sp., and B. subtilis for tomato seed germination and positive effects on plant growth103. Comparative results in the current findings from potential metagenomics clones producing phosphatase with enhanced plant growth promoting properties might be due to the synergistic effect of phosphate solubilising bacteria in different combinations from metagenomic DNA of Arabian Sea sediment. Next-generation sequencing technologies, such as Nanopore, have already been employed to understand the role of plant biostimulants for sustainable agriculture50. In this study, nanopore based long-read metagenomic sequencing of Arabian Sea fosmid clones was utilized to identify the deep-sea microbiome responsible for phosphate solubilisation and to functionally annotate the phosphatase gene potentially promoting plant growth. Recent reports on biofertilizers, especially using phosphate-solubilizing bacteria highlight their ability to enhance nutrient availability, leading to increased crop yields, resilience, and stress tolerance, all of which are crucial for soil health and could help to adapt to changing climate conditions11. The significance of the recent findings aligns with the objectives of current study on unculturable bacterial communities from the Arabian Sea seamount known for their extremophilic properties along with phosphate solubilising ability which could result in promising plant biostimulants with bio-fertilizers application, thereby contributing to sustainable agriculture.
Conclusion
This study highlights the potential use of functional and sequence-based metagenomics for bio-prospecting phosphatase gene clusters from dark sea sediment samples collected from Arabian Sea seamount. About 9068 recombinant clones of metagenome fosmid library was constructed from Arabian Sea seamount sediment which could be a living repository of uncultured deep-sea microbiome thus contributing towards understanding conservation biology of extreme environment and bio-prospecting its functional genes for biotechnological innovations. Nanopore-based metagenomic analysis of five potential clones revealed the presence of phosphate cycling genes hidden in the seamount sediment metagenome which helped to understand the taxonomic diversity and biogeochemical cycling of phosphate solubilising microbes in deep-sea. These recombinant clones that can utilize P sources and secrete phosphatase enzyme along with organic acid production could be a novel approach to enhance soil quality by solubilizing the phosphate in an eco-friendly manner for enhancing Plant growth. This study also highlights the potential of advanced nanopore sequencing techniques to improve agricultural practices by identifying key phosphate cycling genes contributing to soil fertility and plant growth using deep-sea microbes which could be potent plant biostimulants. This approach could pave the way for developing next-generation biofertilizers thereby replacing the chemical fertilisers and restoring the ecosystem health. By leveraging the potential of biotechnological innovations, this study brings an insight to unlock a new era of sustainable agriculture that mitigates climate change and help in addressing global challenges for environmental sustainability using deep-sea microbes.
Statistical analysis
All collected data in this study represent the means of two replicates, which were analysed using one-way analysis of variance (ANOVA). To separate the means, we applied Tukey’s post-hoc test along with a least significant difference (LSD) test (P < 0.05). All statistical analyses were conducted using SPSS version 29.0.
Data availability
The fosmid library sequencing data generated in this study has been submitted to the NCBI Sequence Read Archive (SRA) under the BioProject identifier PRJNA1090364. It can also be accessed through the following link: https://www.ncbi.nlm.nih.gov/sra/PRJNA1090364.
References
-
Sousa, J., Silvério, S. C., Costa, A. M. & Rodrigues, L. R. Metagenomic approaches as a tool to unravel promising biocatalysts from natural resources: soil and water. Catal 12, 385 (2022).
-
Robinson, S. L., Piel, J. & Sunagawa, S. A roadmap for metagenomic enzyme discovery. Nat. Prod. Rep. 38, 1994–2023 (2021).
-
Coutinho, F. H. et al. New viral biogeochemical roles revealed through metagenomic analysis of lake Baikal. Microbiome 8, 163. https://doi.org/10.1186/s40168-020-00936-4 (2020).
-
Sánchez, O., Stefanni, S. & Bhadury, P. The deep sea biodiversity and conservation collection. Sci. Rep. 1, 27559 (2024).
-
Rowden, A. A., Dower, J. F., Schlacher, T. A., Consalvey, M. & Clark, M. R. Paradigms in seamount ecology: fact, fiction and future. Mar. Ecol. 31, 226–241 (2010).
-
Clark, M. R. et al. The ecology of seamounts: structure, function, and human impacts. Ann. Rev. Mar. Sci. 2, 253–278 (2010).
-
Li, H., Zhou, H., Yang, S. & Dai, X. Stochastic and deterministic assembly processes in seamount microbial communities. Appl. Environ. Microbiol. 89, e00701–e00723 (2023).
-
Laiolo, E. et al. Metagenomic probing toward an atlas of the taxonomic and metabolic foundations of the global ocean genome. Front. Sci. 1. https://doi.org/10.3389/fsci.2023.1038696 (2024).
-
Savio, J. J. et al. Morphotectonics of the Raman-Panikkar-Wadia seamount chain in the Laxmi basin, Eastern Arabian sea. J. Asian Earth Sci. 262, 105977 (2024).
-
Savio, J. J. et al. Caldera-like features located over the Panikkar seamount and adjacent regions in the Laxmi basin, Eastern Arabian sea. J. Earth Syst. Sci. 131, 44 (2022).
-
Sreethu, S., Chhabra, V., Kaur, G. & Ali, B. Biofertilizers as a greener alternative for increasing soil fertility and improving food security under climate change condition. Commun. Soil Sci. Plant Anal., 261–285. (2024).
-
Li, J. et al. Quantify the contribution of anammox for enhanced nitrogen removal through metagenomic analysis and mass balance in an anoxic moving bed biofilm reactor. Water Res. 160, 178–187. https://doi.org/10.1016/j.watres.2019.05.070 (2019).
-
Simon, S. A. et al. Dancing the nanopore limbo–nanopore metagenomics from small DNA quantities for bacterial genome reconstruction. Genome 24 (1), 727 (2023).
-
Shum, P., Cusa, M., Prasetyo, A. & Mariani, S. Nanopore sequencing facilitates screening of diversity and provenance of seafood and marine wildlife. Food Control 161, 110382 (2024).
-
Marimuthu, J. et al. Deep-sea sediment metagenome from Bay of Bengal reveals distinct microbial diversity and functional significance. Genomics. 114 (6), 110524 (2022).
-
Balachandran, K. R. S. et al. Microbial community structure and exploration of bioremediation enzymes: functional metagenomics insight into Arabian sea sediments. Mol. Genet. Genomics. 298 (3), 627–651. https://doi.org/10.1007/s00438-023-01995-6 (2023).
-
Fathin, K. P. F. et al. Phosphorus cycling from a coastal upwelling zone in the southeastern Arabian sea. Environ. Monit. Assess. 193, 188. https://doi.org/10.1007/s10661-021-08968-5 (2021).
-
Zeng, J. et al. PCycDB: a comprehensive and accurate database for fast analysis of phosphorus cycling genes. Microbiome 10, 101. https://doi.org/10.1186/s40168-022-01292-1 (2022).
-
Mullaney, E. J. & Ullah, A. H. The term phytase comprises several different classes of enzymes. Biochem. Biophys. Res. Commun. 312, 179–184. https://doi.org/10.1016/j.bbrc.2003.09.176 (2003).
-
Luo, H., Benner, R., Long, R. A. & Hu, J. Subcellular localization of marine bacterial alkaline phosphatases. Proc. Natl. Acad. Sci. U.S.A. 106, 21219–21223. https://doi.org/10.1073/pnas.0907586106 (2009).
-
Sebastian, M. & Ammerman, J. W. The alkaline phosphatase PhoX is more widely distributed in marine bacteria than the classical PhoA. ISME J. 3, 563–572. https://doi.org/10.1038/ismej.2009.10 (2009).
-
Lee, D. H. et al. A novel psychrophilic alkaline phosphatase from the metagenome of tidal flat sediments. BMC Biotechnol. 15, 1 (2015).
-
Crits-Christoph, A., Diamond, S., Butterfield, C. N., Thomas, B. C. & Banfield, J. F. Novel soil bacteria possess diverse genes for secondary metabolite biosynthesis. Nature 558, 440–444. https://doi.org/10.1038/s41586-018-0207-y (2018).
-
Waschulin, V. et al. Biosynthetic potential of uncultured Antarctic soil bacteria revealed through long-read metagenomic sequencing. ISME J. 16, 101–111. https://doi.org/10.1038/s41396-021-01052-3 (2022).
-
Lam, K. N., Cheng, J., Engel, K., Neufeld, J. D. & Charles, T. C. Current and future resources for functional metagenomics. Front. Microbiol. 6. https://doi.org/10.3389/fmicb.2015.01196 (2015).
-
Ren, L. et al. Metagenomics reveals bacterioplankton community adaptation to long-term thermal pollution through the strategy of functional regulation in a subtropical Bay. Water Res. 216, 118298. https://doi.org/10.1016/j.watres.2022.118298 (2022).
-
Batista García, R. et al. From lignocellulosic metagenomes to lignocellulolytic genes: trends, challenges and future prospects: From lignocellulosic metagenomes to lignocellulolytic genes. Biofuel Bioprod. Biorefin. 10. https://doi.org/10.1002/bbb.1709 (2016).
-
Wang, H., Hart, D. & An, Y. Functional metagenomic technologies for the discovery of novel enzymes for biomass degradation and biofuel production. Bioenergy Res. 12. https://doi.org/10.1007/s12155-019-10005-w (2019).
-
Negri, T. et al. A rapid and efficient strategy to identify and recover biosynthetic gene clusters from soil metagenomes. Appl. Microbiol. Biotechnol. 106 (8), 3293–3306. https://doi.org/10.1007/s00253-022-11917-y (2022).
-
Dias, R. et al. Metagenomics: library construction and screening methods. V metagenom. Methods Appl. Perspect. 5, 28–34 (2014).
-
Chuzel, L., Sinha, A., Cunningham, C. V. & Taron, C. H. High-throughput nanopore DNA sequencing of large insert fosmid clones directly from bacterial colonies. Appl. Environ. Microbiol. e0024324. https://doi.org/10.1128/aem.00243-24 (2024).
-
Ngara, T. R. & Zhang, H. Recent advances in function-based metagenomic screening. Genom. Proteom. Bioinform. 16, 405–415 (2018).
-
Selvin, J., Kennedy, J., Lejon, D. P., Kiran, G. S. & Dobson, A. D. Isolation identification and biochemical characterization of a novel halo-tolerant lipase from the metagenome of the marine sponge haliclonasimulans. Microb. Cell. Fact. 11, 72 (2012).
-
Sousa, F. M. O. et al. Functional screening for cellulolytic activity in a metagenomic fosmid library of microorganisms associated with coral. Genet. Mol. Res. 15, 1–8 (2016).
-
Simon, C., Herath, J., Rockstroh, S. & Daniel, R. Rapid identification of genes encoding DNA polymerases by function-based screening of metagenomic libraries derived from glacial ice. Appl. Environ. Microbiol. 75, 2964–2968 (2009).
-
Kim, H., Kwon, K. K., Seong, W. & Lee, S. G. Multi-enzyme screening using a high- throughput genetic enzyme screening system. J. Vis. Exp. 114 (2016).
-
López-López, O., Knapik, K., Cerdán, M. E. & González-Siso, M. I. Metagenomics of an alkaline hot spring in Galicia (Spain): microbial diversity analysis and screening for novel lipolytic enzymes. Front. Microbiol. 6, 1291 (2015).
-
Mohamed, Y. M. et al. Isolation and characterization of a heavy metal-resistant, thermophilic esterase from a red sea Brine pool. Sci. Rep. 3. https://doi.org/10.1038/srep03358 (2013).
-
Nair, H. P. & Bhat, S. G. Arabian sea metagenome derived-α-amylase P109 and its potential applications. Ecol. Genet. Genom. 16, 100060 (2020).
-
Nair, H. P., Vincent, H., Puthusseri, R. M. & Bhat, S. G. Molecular cloning and characterization of a halotolerant α-amylase from marine metagenomic library derived from Arabian sea sediments. Biotech 7, 65. https://doi.org/10.1007/s13205-017-0674-0 (2017).
-
Bhardwaj, Y., Reddy, B. & Dubey, S. K. Organic farming favorsphod-harboringrhizospheric bacterial community and alkaline phosphatase activity in tropical agroecosystem. Plants 12, 1068 (2023).
-
Zheng, M. M., Wang, C., Li, W. X., Song, W. F. & Shen, R. F. Soil nutrients drive function and composition of phoC-harboring bacterial community in acidic soils of Southern China. Front. Microbiol. 10, 2654 (2019).
-
Rizwanuddin, S. et al. Insight into phytase-producing microorganisms for phytate solubilization and soil sustainability. Front. Microbiol. 14, 1127249 (2023).
-
Pan, L. & Cai, B. Phosphate-solubilizing bacteria: advances in their physiology, molecular mechanisms and microbial community effects. Microorganisms 11, 2904 (2023).
-
Compant, S. et al. The plant endosphere world – bacterial life within plants. Environ. Microbiol. 23, 1812–1829. https://doi.org/10.1111/1462-2920.15240 (2021).
-
Hossain, M. M., Sultana, F. & Islam, S. Plant growth-promoting fungi (PGPF): Phytostimulation and induced systemic resistance. In Plant-Microbe Interactions in Agro-Ecological Perspectives: Volume 2: Microbial Interactions and Agro-Ecological Impacts, 135–191. (2017).
-
Mani, G. et al. Halophilic Phosphate-Solubilizing microbes (Priestia megaterium and Bacillus velezensis) isolated from Arabian sea seamount sediments for plant growth promotion. Curr. Microbiol. 81, 1–15 (2024).
-
De Souza, R. S. C., Armanhi, J. S. L. & Arruda, P. From Microbiome to traits: designing synthetic microbial communities for improved crop resiliency. Front. Plant. Sci. 11, 1179 (2020).
-
Marín, O., González, B. & Poupin, M. J. From microbial dynamics to functionality in the rhizosphere: a systematic review of the opportunities with synthetic microbial communities. Front. Plant Sci. 12, 650609. (2021).
-
Garg, S. et al. Next generation plant biostimulants & genome sequencing strategies for sustainable agriculture development. Front. Microbiol. 15, 1439561. https://doi.org/10.3389/fmicb.2024.1439561 (2024).
-
Arora, P. K. et al. Next-generation fertilizers: the impact of bionanofertilizers on sustainable agriculture. Microb. Cell. Fact. 23, 254 (2024).
-
Patin, N. V. & Goodwin, K. D. Capturing marine microbiomes and environmental DNA: A field sampling guide. Front. Microbiol. 13, 1026596 (2023).
-
Mitchell, K. R. & Takacs-Vesbach, C. D. A comparison of methods for total community DNA preservation and extraction from various thermal environments. J. Ind. Microbiol. 35, 1139–1147 (2008).
-
Busi, S. B. et al. Optimised biomolecular extraction for metagenomic analysis of microbial biofilms from high-mountain streams. PeerJ 8, e9973. https://doi.org/10.7717/peerj.9973 (2020).
-
Lam, K. N. et al. Evaluation of a pooled strategy for high-throughput sequencing of cosmid clones from metagenomic libraries. PLoS ONE 9. https://doi.org/10.1371/journal.pone.0098968 (2014).
-
Delaney, S., Murphy, R. & Walsh, F. A comparison of methods for the extraction of plasmids capable of conferring antibiotic resistance in a human pathogen from complex broiler cecal samples. Front. Microbiol. 9, 373868 (2018).
-
Wan, W. et al. Isolation and characterization of phosphorus solubilizing bacteria with multiple phosphorus sources utilizing capability and their potential for lead immobilization in soil. Front. Microbiol. 11, 752 (2020).
-
Thengodkar, R. R. M. & Sivakami, S. Degradation of Chlorpyrifos by an alkaline phosphatase from the Cyanobacterium Spirulina platensis. Biodegradation 21, 637–644. https://doi.org/10.1007/s10532-010-9331-6 (2010).
-
Lindang, H. U., Subbiah, V. K., Rodrigues, K. F. & Budiman, C. Isolation, identification, and characterization of phosphate solubilizing bacteria, Paenibacillus sp., from the soil of Danum Valley tropical rainforest, Sabah. Malaysia Biodiversitas. 22, 4370–4378 (2021).
-
Yu Plisova, E. Y., Balabanova, L. A., Ivanova, E. P., Mikhailov, K. V. B., Agafonova, V. V. & E. V., Rasskazov V. A A highly active alkaline phosphatase from the marine bacterium Cobetia. Mar. Biotechnol. 7, 173–178. https://doi.org/10.1007/s10126-004-3022-4 (2005).
-
Wright, G. S. et al. Purification and structural characterization of aggregation-prone human TDP-43 involved in neurodegenerative diseases. Science 23 (6). (2020).
-
Lowry, O. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193, 265–275 (1951).
-
Weikle, K. Determination of citric acid in fruit juices using HPLC. Concordia Coll. J. Anal. Chem. 3, 57–62 (2012).
-
De Coster, W., D’Hert, S., Schultz, D. T., Cruts, M. & Van Broeckhoven, C. NanoPack: Visualizing and processing long-read sequencing data. Bioinform 34, 2666–2669. https://doi.org/10.1093/bioinformatics/bty149 (2018).
-
Schmieder, R. & Edwards, R. Quality control and pre-processing of metagenomic datasets. Bioinformatics 27, 863–864. https://doi.org/10.1093/bioinformatics/btr026 (2011).
-
Breitwieser, F. P. & Salzberg, S. L. Pavian: Interactive analysis of metagenomics data for microbiomics and pathogen identification. bioRxiv 084715. (2016).
-
Oksanen, J. Vegan: Community ecology package. http://vegan.r-forge.r-project.org/ (2010).
-
Chang Kanehisa, M., Sato, Y., Kawashima, M., Furumichi, M. & Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 44, D457–D462. https://doi.org/10.1093/nar/gkv1070 (2016).
-
Hii, Y. S., Yen San, C. Y., Lau, S. W. & Danquah, M. K. Isolation and characterisation of phosphate solubilizing microorganisms from peat. Biocatal. Agric. Biotechnol. 26, 101643. https://doi.org/10.1016/j.bcab.2020.101643 (2020).
-
Acevedo, E., Galindo-Castañeda, T., Prada, F., Navia, M. & Romero, H. M. Phosphate- solubilizing microorganisms associated with the rhizosphere of oil palm (Elaeis guineensis Jacq.) in Colombia. Appl. Soil. Ecol. 80, 26–33 (2014).
-
Katz, M., Hover, B. M. & Brady, S. F. Culture-independent discovery of natural products from soil metagenomes. J. Ind. Microbiol. 43, 129–141. https://doi.org/10.1007/s10295-015-1706-6 (2016).
-
Daniel, R. The metagenomics of soil. Nat. Rev. Microbiol. 3, 470–478 (2005).
-
Terrón-González, L., Martín-Cabello, G., Ferrer, M. & Santero, E. Functional metagenomics of a biostimulated Petroleum-Contaminated soil reveals an extraordinary diversity of extradiol dioxygenases. Appl. Environ. Microbiol. 82, 2467–2478. https://doi.org/10.1128/AEM.03811-15 (2016).
-
DeLong, E. F. et al. Community genomics among stratified microbial assemblages in the ocean’s interior. Sci 311, 496–503. https://doi.org/10.1126/science.1120250 (2006).
-
Schneegurt, M. A., Dore, S. Y. & Kulpa, C. F. Direct extraction of DNA from soils for studies in microbial ecology. Curr. Issues Mol. Biol. 5, 1–8 (2003).
-
Chen, J. et al. Global marine microbial diversity and its potential in bioprospecting. Nature 1–9. (2024).
-
Ellilä, S. et al. Cloning of novel bacterial Xylanases from lignocellulose-enriched compost metagenomic libraries. AMB Express. 9, 1–12 (2019).
-
Nyyssönen, M. et al. Coupled high-throughput functional screening and next generation sequencing for identification of plant polymer decomposing enzymes in metagenomic libraries. Front. Microbiol. 4, 282. https://doi.org/10.3389/fmicb.2013.00282 (2013).
-
Udaondo, Z. et al. Developing robust protein analysis profiles to identify bacterial acid phosphatases in genomes and metagenomic libraries. Environ. Microbiol. 22, 3561–3571 (2020).
-
Olayide Folashade ObidiOlushina, Awe, O., Okekunjo, M. N. I. E. & F. O Production of phosphatase by microorganisms isolated from discolored painted walls in a typical tropical environment: a Non-Parametric analysis. Arab. J. Basic. Appl. Sci. 25, 111–121. https://doi.org/10.1080/25765299.2018.1527277 (2018).
-
Cheng, Y. et al. Phosphate-solubilizing bacteria: Their agroecological function and optimistic application for enhancing agro-productivity. Sci. Total Environ. 166468. (2023).
-
Behera, B. C. et al. Phosphate solubilization and acid phosphatase activity of Serratia sp. isolated from Mangrove soil of Mahanadi river delta, odisha, India. J. Genet. Eng. &Biotechnol. 15, 169–178 (2017a).
-
Zhang, X. et al. Expression, purification, and characterization of a novel acid phosphatase that displays protein tyrosine phosphatases activity from metarhizium anisopliae strain CQMa102. Biosci. Biotechnol. Biochem. 81, 2292–2300 (2017).
-
Budiman, C., Koga, Y., Takano, K. & Kanaya, S. FK506-binding protein 22 from a psychrophilic bacterium, a cold shock-inducible peptidyl Prolyl isomerase with the ability to assist in protein folding. Int. J. Mol. Sci. 12, 5261–5284 (2011).
-
Razali, R., Subbiah, V. K. & Budiman, C. Technical data of heterologous expression and purification of SARS-CoV-2 proteases using Escherichia coli system. Data 6, 99 (2021).
-
Prasad, P., Kalam, S., Sharma, N. K., Podile, A. R. & Das, S. N. Phosphate solubilization and plant growth promotion by two Pantoea strains isolated from the flowers of Hedychium coronarium L. Front. Agron. 4, 990869 (2022).
-
Carrillo, A. E., Li, C. Y. & Bashan, Y. Increased acidification in the rhizosphere of cactus seedlings induced by azospirillumbrasilense. Sci. Nat. 89, 428–432 (2002).
-
Puente, M. E., Bashan, Y., Li, C. Y. & Lebsky, V. K. Microbial populations and activities in the rhizoplane of rock- weathering desert plants. I. Root colonization and weathering of igneous rocks. Plant. Biol. 6, 629–642 (2004).
-
Lal, L. Phosphate biofertilizers. (Agrotech Publ. Academy, 2002).
-
Behera, B. C. et al. Alkaline phosphatase activity of a phosphate solubilizing Alcaligenes faecalis, isolated from Mangrove soil. Biotechnol. Res. Innov. 1, 101–111. https://doi.org/10.1016/j.biori.2017.01.003 (2017b).
-
Mardad, I., Serrano Delgado, A. & Soukri, A. Solubilization of inorganic phosphate and production of organic acids by bacteria isolated from a Moroccan mineral phosphate deposit. Afr. J. Microbiol. Res. 7, 626–635 (2013).
-
Chang, H. M., Foo, H. L., Loh, T. C., Lim, E. T. C., Mutalib, A. & N. E Comparative studies of inhibitory and antioxidant activities, and organic acids compositions of postbiotics produced by probiotic Lactiplantibacillus plantarum strains isolated from Malaysian foods. Front. Vet. Sci. 7, 602280 (2021).
-
Ding, L. et al. Genome-wide analysis ofBES1/BZR1 transcription factors and their responses to osmotic stress in Ammopiptanthus nanus. J. Res. 26, 127–135 (2021).
-
Stevens, B. M., Creed, T. B., Reardon, C. L. & Manter, D. K. Comparison of Oxford Nanopore Technologies and Illumina MiSeq sequencing with mock communities and agricultural soil. Sci. Rep. 13 (1), 9323 (2023).
-
Das, S., Lyla, P. S. & Khan, S. A. Biogeochemical processes in the continental slope of Bay of bengal: I. Bacterial solubilization of inorganic phosphate. Rev. Biol. Trop. 55, 1–9. https://doi.org/10.15517/rbt.v55i1.6052 (2007).
-
Paul, D. & Sinha, S. N. Isolation and characterization of phosphate solubilizing bacterium Pseudomonas aeruginosa KUPSB12 with antibacterial potential from river ganga, India. Ann. Agrar. Sci. 15, 130–136 (2017).
-
Gupta, R. et al. Identification, characterization and optimization of phosphate solubilizing rhizobacteria(PSRB) from rice rhizosphere. Saudi J. Biol. Sci. 29, 35–42. https://doi.org/10.1016/j.sjbs.2021.09.075 (2022).
-
Khatoon, N. & Khan, M. Evaluation of Bacillus subtilis MRB4, as plant growth promoter and potential phosphate solubilizer under abiotic stress. J. Appl. Biol. Biotechnol. 8, 27–35 (2020).
-
Monds, R. D., Newell, P. D., Schwartzman, J. A. & O’Toole, G. A. Conservation of the Pho Regulon in Pseudomonas fluorescens Pf0-1. Appl. Environ. Microbiol. 72, 1910–1924. https://doi.org/10.1128/AEM.72.3.1910-1924.2006 (2006).
-
Su, Y., Liu, C., Fang, H. & Zhang, D. Bacillus subtilis: a universal cell factory for industry, agriculture, biomaterials and medicine. Microb. Cell. Fact. 19, 1–12 (2020).
-
Nawaz, M. Z., Subin Sasidharan, R., Alghamdi, H. A. & Dang, H. Understanding interaction patterns within deep-sea microbial communities and their potential applications. Mar. Drugs. 20, 108 (2022).
-
Pappalettere, L., Bartolini, S. & Toffanin, A. Enhancement of tomato seed germination and growth parameters through seed priming with Auxin-Producing plant growth promoting Bacteria strains. Seeds 3, 479–492 (2024).
-
Kumar, P. et al. Seed bio-priming with tri-species consortia of phosphate solubilizing rhizobacteria (PSR) and its effect on plant growth promotion. Heliyon 6, 12 (2020).
Acknowledgements
The authors are grateful to the Director, National Institute of Ocean Technology, for providing the necessary facilities to carry out this study. The authors acknowledge the funding received from Ministry of Earth Sciences, Government of India under Deep Ocean Mission Program for carrying out this research work. The authors also express their gratitude to the NIOT Vessel Management Cell and crew members of ORV Sagar Manjusha (SAMA10) for their help during the collection of deep-sea sediment samples. We are thankful to Genotypic Technology Pvt. Ltd., Bangalore, India for Nanopore sequencing. We express our gratitude to Dr. M. Savurirajan, Project Scientist I, and Dr. Dilip Kumar Jha, Scientist-F, Marine Biotechnology division, National Institute of Ocean Technology, Ministry of Earth Sciences, Government of India, Chennai 600100, India, for preparing bathymetric mapping for the sample collected.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Balachandran, K., Mani, G., Sidharthan, A.T. et al. Unearthing the genetic resources of Arabian sea seamount and metagenomic insights into phosphate cycling genes for next generation plant biostimulants. Sci Rep 15, 35782 (2025). https://doi.org/10.1038/s41598-025-10337-y
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-10337-y